Вторично активный транспорт. Котранспорт глюкозы и аминокислот в клетке
Добавил пользователь Дмитрий К. Обновлено: 21.01.2026
Презентация на тему: " Вторично- активный транспорт через мембрану. введение Что такой вторичный активный транспорт? Он заключается в транспорте вещества против градиента,обеспечиваемом." — Транскрипт:
1 Вторично- активный транспорт через мембрану
2 введение Что такой вторичный активный транспорт? Он заключается в транспорте вещества против градиента,обеспечиваемом энергией,которая высвобождается при транспорте другого вещества по градиенту. Если же в качестве источника энергии используется градиент концентрации ионов,то транспорт называют вторично-активным.
3 Вторичный активный транспорт - чрезвычайно важный для растительной клетки процесс. Благодаря ему клетка активно поглощает (или удаляет) многие вещества (ионы, углеводы, аминокислоты и др.). Суть этого процесса состоит в том, что в мембране имеются особые вещества - переносчики белковой природы. Они могут образовывать комплекс с протоном на наружной стороне мембраны. Такой комплекс приобретает сродство (в зависимости от типа переносчика) к определенному веществу (например, иону Na+, сахарозе) на одной из сторон мембраны и образуется заряженное соединение типа протон- переносчик-вещество. Перенося протон внутрь клетки как по электрическому (Dy), так и по концентрационному (DрН) градиентам, переносчик за счет энергии этих двух составляющих протонного потенциала переносит вещество внутрь (симпорт) или наружу (антипорт)
4 Симпорт (котранспорт) Симпорт также использует процесс диффузии. В этом случае молекула, которая всегда стремиться в клетку по пути диффузии, используется для того, чтобы перетащить за собой другую молекулу. В данном примере глюкоза проникает в клетку вселд за натрием Симпорт это активный перенос вещества через мембрану, осуществляемый за счет энергии градиента концентрации другого вещества.
5 снаружи мембрана внутри аминокислота Na + переносчик Гипотетический механизм опосредованного натрием котранспорта аминокислот(или сахаров ). Для осуществления транспорта переносчик должен связать и Na +,и аминокислоту.Направленный внутрь транспорт осуществляется за счет градиента Na +.
6 мембрана мембрана Метаболический насос АТР АДР + Рi Na + Общий переносчик Na + Аминок-ислота сахар Na + Аминокис-лота сахар Транспорт аминокислот и сахаров зависит в конечном счете от химической энергии,запасенной в АТР в процессе клеточного метаболизма.Концентрационный градиент Na+ можно рассматривать как некую промежуточную форму потенциальной энергии,используемую для перемещения органических молегул против их концентрационных градиентов.
7 Антипорт (контртранспорт) Антипорт - это перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту своей концентрации. Антипорт - это перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту своей концентрации.
8 Некоторые белки функционируют как котранспортные системы, в которых перенос одного растворенного вещества зависит от одновременного или последовательного переноса другого вещества либо в том же направлении, либо в противоположном Перенос другого вещества в противоположном направлении называется антипорт. Например, натрий-калиевый насос в эукариотических плазматических мембранах работает по принципу антипорта, качая ионы натрия из клетки, а ионы калия - внутрь клетки.
10 вывод Симпорт и антипорт могут происходить при всасывании аминокислот из кишечника и реабсорбции глюкозы из первичной мочи. При этом используется энергия градиента концентрации ионов Na+, создаваемого Na+, K+-АТР-азой. Симпорт и антипорт могут происходить при всасывании аминокислот из кишечника и реабсорбции глюкозы из первичной мочи. При этом используется энергия градиента концентрации ионов Na+, создаваемого Na+, K+-АТР-азой.
Транспорт через мембрану.
Внутриклеточные органеллы также могут обладать избирательно проницаемой мембраной. Например, в лизосомах мембрана поддерживает концентрацию ионов водорода (Н + ) в 1000-10000 раз больше, чем в цитозоле.
Транспорт через мембрану может быть пассивным, облегченным или активным.
1.1. Пассивный транспорт - это движение молекул или ионов по концентрационному либо электрохимическому градиенту. Это может быть простая диффузия, как в случае проникновения через плазматическую мембрану газов (например О2 и СО2 ) или простых молекул (этанола). При простой диффузии растворенные во внеклеточной жидкости небольшие молекулы последовательно растворяются в мембране и затем во внутриклеточной жидкости. Указанный процесс неспецифичен, при этом скорость проникновения через мембрану определяется степенью гидрофобности молекулы, то есть ее жирорастворимостью. Скорость диффузии через липидный бислой прямо пропорциональна гидрофобности, а также трансмембранному градиенту концентрации или электрохимическому градиенту.
1.2. Облегченная диффузия - это быстрое движение молекул через мембрану с помощью специфических мембранных белков, называемых пермеазами. Этот процесс специфичен, он протекает быстрее простой диффузии, но имеет ограничение скорости транспорта.
Облегченная диффузия обычно характерна для водорастворимых веществ. Большинство (если не все) мембранных переносчиков являются белками. Конкретный механизм функционирования переносчиков при облегченной диффузии исследован недостаточно. Они могут, например, обеспечивать перенос путем вращательного движения в мембране. В последнее время появились сведения, что белки-переносчики при контакте с транспортируемым веществом изменяют свою конформацию, в результате в мембране открываются своеобразные «ворота», или каналы. Эти изменения происходят за счет энергии, высвобождающейся при связывании транспортируемого вещества с белком. Возможен также перенос эстафетного типа. В этом случае сам переносчик остается неподвижным, а ионы мигрируют вдоль него от одной гидрофильной связи к другой.
Моделью переносчика такого типа может служить антибиотик грамицидин. В липидном слое мембраны его длинная линейная молекула принимает форму спирали и образует гидрофильный канал, по которому может мигрировать по градиенту ион К.
Получены экспериментальные доказательства существования природных каналов в биологических мембранах. Транспортные белки отличаются высокой специфичностью по отношению к переносимому через мембрану веществу, по многим свойствам напоминая ферменты. Они обнаруживают большую чувствительность к рН, конкурентно ингибируются соединениями, близкими по структуре к переносимому веществу, и неконкурентно - агентами, изменяющими специфически функциональные группы белков.
Облегченная диффузия отличается от обычной не только скоростью, но и способностью к насыщению. Увеличение скорости переноса веществ происходит пропорционально росту градиента концентрации только до определенных пределов. Последний определяется «мощностью» переносчика.
1.3. Активный транспорт - это движение ионов или молекул через мембрану против градиента концентрации за счет энергии гидролиза АТФ. Имеются три основных типа активного транспорта ионов:
1. натрий-калиевый насос - Na + /K + -аденозинтрифосфатаза (АТФаза), переносящая Na + наружу, а K + внутрь;
2. кальциевый (Са 2+ ) насос - Са 2+ -АТФаза, которая транспортирует Са 2+ из клетки или цитозоля в саркоплазматический ретикулум;
3. протонный насос - Н + -АТФаза. Созданные активным транспортом градиенты ионов могут быть использованы для активного транспорта других молекул - таких, как некоторые аминокислоты и сахара (вторичный активный транспорт).
Котранспорт - это транспорт иона или молекулы, сопряженный с переносом другого иона. Симпорт - одновременный перенос обеих молекул в одном направлении; антипорт - одновременный перенос обеих молекул в противоположных направлениях. Если транспорт не сопряжен с переносом другого иона, этот процесс называется унипортом. Котранспорт возможен как при облегченной диффузии, так и в процессе активного транспорта.
Глюкоза может транспортироваться путем облегченной диффузии по типу симпорта. Ионы Cl - и HCO3 - транспортируются через мембрану эритроцитов путем облегченной диффузии переносчиком, называемым полосой 3, по типу антипорта. При этом Cl - и HCO3 - переносятся в противоположных направлениях, а направление переноса определяется преобладающим градиентом концентрации.
Активный транспорт ионов против градиента концентрации требует энергии, выделяемой при гидролизе АТФ до АДФ: АТФ à АДФ + Ф (неорганический фосфат). Для активного транспорта, как и для облегченной диффузии, характерны: специфичность, ограничение максимальной скорости (то есть кинетическая кривая выходит на плато) и наличие ингибиторов. В качестве примера можно привести первичный активный транспорт, осуществляемый Na + /K + - АТФазой. Для функционирования этой фрментной системы антипорта необходимо наличие Na + , K + и ионов магния. Она присутствует практически во всех клетках животных, причем ее концентрация особенно высока в возбудимых тканях (например, в нервах и мышцах) и в клетках, принимающих активное участие в движении осуществляемый Na + через плазматическую мембрану (например, в корковом слое почек и слюнных железах).
Сам фермент АТФаза представляет собой олигомер, состоящий из 2 a-субъедениц по 110 кД и 2 гликопротеиновых b-субъдениц по 55 кД каждая.. при гидролизе АТФ происходит обратимое фосфорилирование определенного остатка аспартата на a-субъеденице с образованием b-аспартамилфосфата.. Для фосфорилирования необходимы Na + и Мg 2+ , но не K + , тогда как для дефосфорилирования необходим K + , но не Na + или Мg 2+ . Описаны два конформационных состояния белкового комплекса с различным энергетическим уровнем, которые принято обозначать Е1 и Е2 , поэтому АТФазу называют также переносчиком типа Е1 - Е2 . Сердечные гликозиды, например дигоксин и уабаин, подавляют активность АТФазы.. Уабаин вследствие хорпошой растворимости в воде широко применяют в экспериментальных исследованиях для изучения натриевого насоса.
Общепринятое представлени о работе Na + /K + - АТФазой , сводится к следующему. Ионы Na и АТФ присоединяются к молекуле АТФазы в присутствии Мg 2+ . Связывание ионов Na запускает реакцию гидролиза АТФ, в результате которой образуются АДФ и фосфорилированная форма фермента. Фосфорилирование индуцирует переход ферментативного белка в новое конформационное состояние и участок или участки, несущие Na, оказываются обращенными к внешней среде. Здесь Na + обменивается на K + , так как для фосфорилированной формы ферментахарактерно высокое сродство к ионам К. обратный переход фермента в исходную конформацию инициируется гидролитическим отщеплением фосфорильной группы в виде неорганического фосфата и сопровождается освобождением K + во внутреннее пространство клетки. Дефосфорилированный активный центр фермента способен присоединить новую молекулу АТФ, и цикл повторяется.
Количества поступивших в клетку в результате работы насоса ионов К и Na не равны между собой. На три выведенных иона Na приходится два введенных иона К при одновременном гидролизе одной молекулы АТФ. Открывание и закрывание канала на противоположных сторонах мембраны и чередующееся изменение эффективности связывания Na и К обеспечиваются энергией гидролиза АТФ. Транспортируемые ионы - Na и К - кофакторы данной ферментативной реакции. Теоретически можно представить самые различные насосы, действующие по этому принципу, хотя в настоящее время известны лишь немногие из них.
1.4. Транспорт глюкозы.Транспорт глюкозы может происходить по типу как облегченной диффузии, так и активного транспорта, причем в первом случае он протекает как унипорт, во втором - как симпорт. Глюкоза может транспортироваться в эритроциты путем облегченной диффузии. Константа Михаэлиса (Кm)для транспорта глюкозы в эритроциты составляет приблизительно 1,5 ммоль/л (то есть при этой концентрации глюкозы около 50% имеющихся молекул пермеазы будет связано с молекулами глюкозы). Поскольку концентрация глюкозы в крови человека составляет 4-6 ммоль/л, поглощение ее эритроцитами происходит практически с максимальной скоростью. Специфичность пермеазы проявляется уже в том, что L-изомер почти не транспортируется в эритроциты в отличие от D-галактозы и D-маннозы, но для достижения полунасыщения транспортной системы требуются более высокие их концентрации. Оказавшись внутри клетки, глюкоза подвергается фосфорилированию и более не способна покинуть клетку. Пермеазу для глюкозы называют также D-гексозной пермеазой. Она представляет собой интегральный мембранный белок с молекулярной массой 45кД.
Глюкоза может также транспортироваться Na + -зависимой системой симпорта, обнаруженной в плазматических мембранах ряда тканей, в том числе в канальцах почек и эпителии кишечника. При этом одна молекула глюкозы переносится путем облегченной диффузии против градиента концентрации, а один ион Na - по градиенту концентрации. Вся система в конечном счете функционирует за счет насосной функции Na + /K + - АТФазы. Таким образом, симпорт является вторичной системой активного транспорта. Аминокислоты транспортируются аналогичным образом.
1.5. Ca 2+ -насоспредставляет собой систему активного транспорта типа Е1 - Е2 , состоящую из интегрального мембранного белка, который в процессе переноса Ca 2+ фосфорилируется по остатку аспартата. При гидролизе каждой молекулы АТФ происходит перенос двух ионов Ca 2+ . В эукариотических клетках Ca 2+ может связываться с кальцийсвязывающим белком, называемым кальмодулином, и весь комплекс связывается с Ca 2+ -насосом. К Ca 2+ -связывающим белкам отнсятся также тропонин С и парвальбумин.
Ионы Са, подобно ионам Na, активно выводятся из клеток Ca 2+ -АТФазой. Особенно большое количество белка кальциевого насоса содержат мембраны эндоплазматического ретикулума. Цепь химических реакций, ведущих к гидролизу АТФ и перебросу Ca 2+ , может быть записана в виде следующих уравнений:
Где Сан - Ca 2+ , находящийся снаружи;
Савн - Ca 2+ , находящийся внутри;
Е1 и Е2 - различные конформации фермента переносчика, переход которых из одной в другую связан с использованием энергии АТФ.
Система активного вывода Н + из цитоплазмы поддерживается двумя типами реакций: деятельностью электрон-транспортной цепи (редокс-цепи) и гидролизом АТФ. Оба - и редокс- и гидролитический Н + -насосы - находятся в мембранах, способных превращать световую или химическую энергию в энергию DmН + (то есть плазматических мембранах прокариот, сопрягающих мембранах хлоропластов и митохондрий). В результате работы Н + АТФазы и/или редокс-цепи транслоцируются протоны, и на мембране возникает протондвижущая сила (DmН + ). Электрохимический градиент ионов водорода, как показывают исследования, может быть использован для сопряженного транспорта (вторичный активный транспорт) большого числа метаболитов - анионов, аминокислот, сахаров и т.д.
С активностью плазматической мембраны связаны обеспечивающие поглощение клеткой твердых и жидких веществ с большой молекулярной массой, - фагоцитоз и пиноцитоз (от герч. фагос - есть, пинос - пить, цитос - клетка). Клеточная мембрана образует карманы, или впячивания, которые втягивают вещества извне. Затем такие впячивания отшнуровываются и окружают мембраной капельку внешней среды (пиноцитоз) или твердые частицы (фагоцитоз). Пиноцитоз наблюдается в самых разнообразных клетках, особенно в тех органах, где происходят процессы всасывания.
Вторично-активный транспорт(перенос аминокислот и глюкозы в энтероцитах и в почечных канальцах)
3) В результате градиента концентрации (для натрия) этот переносчик + глюкоза + ион натрия переносится внутрь клетки, где глюкоза и натрий отщепляются от переносчика, а переносчик «уходит» вновь совершать свою работу.
4) Натрий откачивается помпой, а глюкоза покидает клетку с другой стороны уже по градиенту концентраций (облегченная диффузия по типу «перенос с участием переносчика»).
Котранспорт - вид транспорта, когда одним механизмом транспортируется одновременно две молекулы.
В случае калиево-натриевого насоса имеет место антипорт - транспорт ионов в противоположных направлениях за счет электрохимического градиента одного из ионов (калий идет в одну сторону, натрий - в противоположную),
В случае вторично-активного транспорта - симпорт - транспорт ионов в одном направлении за счет электрохимического градиента одного из ионов.
Показатели характеризующие возбудимость , проводимость и лабильность
Показатели возбудимости:
Порог раздражения - минимальная сила раздражителя, способная вызвать возбуждение.
Реобаза - минимальная сила тока, вызывающая возбуждение.
Хронаксия - минимальное время действия раздражителя силой в 2 реобазы необходимое для возникновения ПД.
Длительность абсолютной рефрактерной фазы.
Показатели проводимости:
Скорость проведения ПД( у нерва 120 м/c).
Показатели лабильности:
Максимальное число импульсов в 1 с.
Методы определения лабильности возбудимых тканей
Лабильность определяется длительностью фазы абсолютной рефрактерности - Наносится надпороговый стимул, который генерирует ПД, затем через интервал подается второй стимул той же силы. Если он вызовет ПД, то сокращают интервал до тех пор, пока даже увеличенный второй стимул не сможет вызвать ПД.
Определяется по максимальному числу возбуждений в единицу времени в ответ на ритмические раздражения.(У нерва 1000 импульсов в сек. У мышц 500 - 250 импульсов в сек.)
Методы исследования возбудимости нервов и мышц. Хронаксиметрия.
Для исследования возбудимости применяют порог раздражения, реобазу, хронаксию, скорость аккомодации, минимальный градиент, длительность АРФ и ОРФ(абсол. и относ. рефрактерность).
Хронаксиметрия— методика измерения хронаксии для исследования возбудимости живых тканей с учетом не только силы раздражителя, но и фактора времени — длительности действия раздражителя.
1)индифферентный электрод хронаксиметра фиксируют на предплечье испытуемого.
2)Переключатель хронаксиметра устанавливают для измерения реобазы (пороговое напряжение тока), при этом хронаксиметр генерирует импульсы тока большей длительности.
3) При напряжении тока 20-40 В находят точку, раздражение которой активным электродом вызывает сокращение мышцы. Отмечают эту точку химическим карандашом. Переводят реостат в нулевое положение.
4) Устанавливают активный электрод на отмеченную карандашом точку. Регистрируя реостатом напряжение тока, находят величину порогового раздражения.
5)Переводят переключатель с измерения реобазы на измерение хронаксии и удваивают напряжение тока. Начиная с минимальных величин, увеличивают длительность импульсов до появления сокращения мышцы и определяют хронаксию
I.10. Транспорт веществ в клетке
Вещества циркулируют в клетке, будучи упакованными в мембраны («передвижение содержимого клетки в контейнерах»). Сортировка веществ и их передвижение связаны с наличием в мембранах комплекса Гольджи специальных белков-рецепторов. Транспорт через мембраны, в том числе и через плазматическую мембрану (цитолемму), является одной из важнейших функций живых клеток. Различают два типа транспорта: пассивный и активный. Пассивный транспорт не требует затрат энергии, активный транспорт энергозависимый. Пассивный транспорт незаряженных молекул осуществляется по градиенту концентрации путем диффузии. Транспорт заряженных веществ зависит от разности потенциалов на поверхности цитолеммы. Как правило, внутренняя цитоплазматическая поверхность мембраны несет отрицательный заряд, что облегчает проникновение в клетку положительно заряженных ионов. Переход ионов или молекул из зоны, где эти вещества находятся в более высокой концентрации, в зону с более низкой концентрацией получил название диффузии. Специфические транспортные белки, встроенные в мембрану, переносят через нее небольшие молекулы. Каждый транспортный белок осуществляет транспорт одного класса молекул или только одного соединения. Трансмембранные белки либо являются переносчиками, либо образуют «каналы». Диффузия может быть нейтральной, когда незаряженные вещества проходят между липидными молекулами или через белки цитолеммы, формирующие каналы. «Облегченная» диффузия происходит при участии специфических белков-переносчиков, связывающих вещество и переносящих его через мембрану. «Облегченная» диффузия протекает быстрее, чем нейтральная. Активный транспорт осуществляют белки-переносчики. При этом расходуется энергия, возникающая вследствие гидролиза АТФ, а также разных потенциалов (зарядов) на различных поверхностях мембраны. Активный транспорт происходит против градиента концентрации. На цитолемме с помощью натриево-калиевого насоса поддерживается мембранный потенциал. Этот насос, накачивающий ионы К+ в клетку против градиентов концентрации, а ионы Na+ - во внеклеточное пространство, является ферментом АТФазой. Благодаря АТФазе ионы Na+ переносятся через мембрану и выводятся во внеклеточную среду, а ионы К+ переносятся внутрь клетки. АТФаза осуществляет также активный транспорт аминокислот и сахаров. Активный транспорт веществ через мембраны. Натрий-калиевый насос. Иногда необходимо, чтобы внутри клетки концентрация вещества была высокой даже при низкой концентрации его во внеклеточной жидкости (например, для ионов калия). И наоборот, концентрацию других ионов внутри клетки важно сохранять на низком уровне, несмотря на их высокие концентрации вне клетки (например, для ионов натрия). Ни в одном из этих двух случаев это не может обеспечиваться простой диффузией, итогом которой всегда является уравновешивание концентрации ионов по обе стороны мембраны. Для создания избыточного движения ионов калия внутрь клетки, а ионов натрия — наружу необходим некий источник энергии. Процесс перемещения молекул или ионов через клеточную мембрану против градиента концентрации (или против электрического градиента, а также градиента давления) называют активным транспортом. К веществам, активно транспортируемым, по крайней мере, через некоторые клеточные мембраны, относят ионы натрия, калия, кальция, железа, водорода, хлора, йода, мочевой кислоты, некоторые сахара и большинство аминокислот. Первично активный и вторично активный транспорт. В зависимости от источника используемой энергии активный транспорт подразделяется на два типа: первично активный и вторично активный. Для первично активного транспорта энергия извлекается непосредственно при расщеплении аденозинтрифосфата или некоторых других высокоэнергетических фосфатных соединений. Вторично активный транспорт обеспечивается вторичной энергией, накопленной в форме разности концентраций побочных веществ, молекул или ионов, по обе стороны клеточной мембраны, созданной первоначально первично активным транспортом. В обоих случаях, как и при облегченной диффузии, транспорт зависит от белков-переносчиков, пронизывающих клеточную мембрану. Однако функции белков-переносчиков при активном транспорте отличаются от переноса облегченной диффузией, поскольку в первом случае белки способны передавать энергию транспортируемому веществу для его перемещения против электрохимического градиента. Далее приведены примеры первично активного и вторично активного транспорта с более детальными объяснениями принципов их функционирования. (рис. 4)
![]()
Натрий-калиевый насос. К веществам, которые транспортируются посредством первично активного транспорта, относят натрий, калий, кальций, водород, хлор и некоторые другие ионы. Механизм активного транспорта лучше всего изучен для натрий-калиевого насоса (Na+/K+-нaсоса) — транспортного процесса, который выкачивает ионы натрия через мембрану клетки наружу и в то же время закачивает в клетку ионы калия. Этот насос отвечает за поддержание различной концентрации ионов натрия и калия по обе стороны мембраны, а также за наличие отрицательного электрического потенциала внутри клеток. (В главе 5 будет показано, что он является также основой процесса передачи импульсов в нервной системе.)
Белок-переносчик представлен комплексом из двух раздельных глобулярных белков: более крупного, называемого альфа-субъединицей, с молекулярной массой около 100000, и меньшего, называемого бета-субъединицей, с молекулярной массой около 55000. Хотя функция меньшего белка неизвестна (за исключением того, что он, возможно, закрепляет белковый комплекс в липидной мембране), крупный белок имеет три специфических характеристики, важные для функционирования насоса. 1. На выступающей внутрь клетки части белка имеются три рецепторных участка для связывания ионов натрия. 2. На наружной части белка располагаются два рецепторных участка для связывания ионов калия. 3. Внутренняя часть белка, расположенная вблизи участков связывания ионов натрия, обладает АТФ-азной активностью. Рассмотрим работу насоса. Когда 2 иона калия связываются с белком-переносчиком снаружи и 3 иона натрия связываются с ним внутри, активируется АТФ-азная функция белка. Это ведет к расщеплению 1 молекулы АТФ до АДФ с выделением энергии высокоэнергетической фосфатной связи. Полагают, что эта освобожденная энергия вызывает химическое и конформационное изменение молекулы белка-переносчика, в результате 3 иона натрия перемещаются наружу, а 2 иона калия — внутрь клетки. Как и другие ферменты, Na-K+-ATФ-aзa может работать и в обратном направлении. При экспериментальном увеличении электрохимических градиентов для Na+ и К+ до таких значений, что накопленная в них энергия станет выше химической энергии гидролиза АТФ, эти ионы будут двигаться по своим градиентам концентрации, а Na+/K+-Hacoc будет синтезировать АТФ из АДФ и фосфата. Следовательно, фосфорилированная форма Nа+/К+ -насоса может быть или донором фосфатов для синтеза АТФ из АДФ, или использовать энергию для изменения своей конформации и качать натрий из клетки, а калий — в клетку. Относительные концентрации АТФ, АДФ и фосфатов, как и электрохимические градиенты для натрия и калия, определяют направление ферментативной реакции. Для некоторых клеток, например, электрически активных нервных клеток, от 60 до 70% всей потребляемой клеткой энергии тратится на перемещение натрия наружу и калия внутрь. Роль Na-K-насоса: Активный транспорт ионов кальция и водорода в клетке. Одной из многочисленных важных функций Na+/K+-Hacoca является регуляция объема каждой клетки. Без функционирования этого насоса большинство клеток организма будут набухать, пока не лопнут. Механизм регуляции объема следующий: внутри клетки много белков и других органических молекул, которые не могут покинуть клетку. Большинство из них отрицательно заряжены и потому связывают большое число ионов калия, натрия и других положительно заряженных ионов. Все эти молекулы и ионы вызывают осмос воды в клетку. Без регуляции осмоса клетка будет неограниченно разбухать вплоть до разрыва мембраны. В норме механизмом для предупреждения этого является Na+/K+-Hacoc. Вспомним, что в результате работы насоса 3 иона натрия выводятся наружу, а 2 иона калия закачиваются внутрь. Кроме того, мембрана гораздо менее проницаема для ионов натрия, чем для калия, поэтому ионы натрия, оказавшись снаружи, в основном там и остаются. Следовательно, присутствует общая потеря ионов клеткой, что, в свою очередь, инициирует осмос воды из клетки. Когда клетка начинает разбухать, это автоматически активирует Na+/K+-Hacoc, обеспечивая удаление из клетки еще большего числа ионов вместе с водой. Таким образом, Na+/K+-Hacoc осуществляет непрерывную регуляцию объема клетки, поддерживая его в нормальных пределах. Электрогенная природа натрий-калиевого насоса. Как известно, Na+/K+-Hacoc выкачивает 3 иона натрия наружу на каждые 2 иона калия, входящие внутрь. Это означает, что 1 положительный заряд выводится наружу при каждом цикле работы насоса. Создается избыток положительных зарядов на поверхности клетки и дефицит положительных ионов внутри клетки, т.е. внутренняя часть клетки заряжается отрицательно. В связи с этим Na+/K+-Hacoc называют электрогенным, поскольку он создает трансмембранную разность потенциалов, а наличие электрического потенциала является основой для передачи сигналов в нервных и мышечных волокнах. Первично активный транспорт ионов кальция. Другим важным механизмом первично активного транспорта является кальциевый насос. В норме ионы кальция во внутриклеточном цитозоле практически всех клеток содержатся в чрезвычайно низкой концентрации — примерно в 10000 раз меньшей, чем во внеклеточной жидкости. Это обеспечивается главным образом двумя кальциевыми насосами. Один из них находится в клеточной мембране и выкачивает ионы кальция из клетки. Другой перекачивает ионы кальция в одну или более ячеистых внутриклеточных органелл, таких как саркоплазматический ретикулум в мышечных клетках или митохондрии во всех клетках. В каждом из этих случаев белок-переносчик пронизывает мембрану насквозь и функционирует как АТФ-аза, имеющая такую же способность расщеплять АТФ, как и АТФ-аза белка-переносчика ионов натрия. Различие заключается в том, что этот белок имеет высокоспецифическое место связи для кальция, а не для натрия. Первично активный транспорт ионов водорода особенно важен в двух участках тела: (1) в железах желудка; (2) в концевой части дистальных канальцев и кортикальных отделах собирательных трубочек почек. (рис.5)
![]()
В железах желудка глубоко расположенные париетальные клетки имеют самый мощный первично активный механизм для переноса ионов водорода по сравнению с любой другой частью тела. Это является основой для секреции соляной кислоты в желудке. В секреторных концах париетальных клеток желудочных желез концентрация ионов водорода повышается в миллион раз, затем они выделяются в желудок вместе с ионами хлора, формируя соляную кислоту. В почечных канальцах имеются специальные вставочные клетки в концевой части дистальных канальцев и в кортикальных отделах собирательных трубочек, где также происходит первично активный транспорт ионов водорода. В этом случае большое количество ионов водорода секретируется из крови в мочу с целью удаления избытка этих ионов из жидкостей организма против градиента концентрации, примерно в 900 раз. Количество энергии, необходимое для активного переноса вещества через мембрану, определяется степенью концентрации вещества во время переноса. Так, 100-кратное концентрирование требует энергии в 2 раза больше по сравнению с энергией, необходимой для увеличения концентрации вещества в 10 раз, а для 1000-кратного концентрирования энергии нужно в 3 раза больше. Другими словами, необходимая энергия пропорциональна десятичному логарифму степени концентрирования вещества и выражается следующей формулой: Энергия (кал/осм) = 1400 log(C1/C2) Для концентрирования 1 осмоля вещества в 10 раз нужно примерно 1400 калорий, а для концентрирования в 100 раз — 2800 калорий. Очевидно, что энергия, расходуемая для концентрирования веществ в клетках или для удаления веществ из клеток против градиента концентрации, должна быть огромной. Некоторые клетки, например выстилающие почечные канальцы и многие железистые клетки, только на эти цели тратят до 90% своей энергии. Вторично активный транспорт. Котранспорт глюкозы и аминокислот в клетке. Когда ионы натрия транспортируются наружу клетки с помощью первично активного транспорта, обычно возникает высокий концентрационный градиент ионов натрия: высокая концентрация снаружи и очень низкая концентрация внутри клетки. Этот градиент является хранилищем энергии, поскольку избыток натрия снаружи клеточной мембраны всегда пытается диффундировать внутрь клетки. При соответствующих условиях эта энергия диффузии натрия может перемещать через мембрану вместе с натрием другие вещества. Это явление называют котранспортом, оно представляет одну из форм вторично активного транспорта. Для транспорта вместе с ионами натрия другого вещества необходим механизм сопряжения. Это обеспечивается с помощью еще одного белка-переносчика в клеточной мембране. Переносчик в этом случае служит местом прикрепления, как для иона натрия, так и для транспортируемого в этом же направлении другого вещества. Когда они оба прикрепляются к белку, энергетический градиент иона натрия обеспечивает совместный перенос иона натрия и другого вещества внутрь клетки. При контртранспорте ионы натрия также пытаются диффундировать внутрь клетки в связи с их высоким градиентом концентрации. Однако на этот раз транспортируемое вещество находится внутри клетки и должно быть выведено наружу. Следовательно, ион натрия связывается с участком белка-переносчика на наружной стороне мембраны, в то время как транспортируемое в обратном направлении вещество связывается с выступающим внутрь клетки участком этого белка. Сразу после их связывания происходят конформационные изменения, и энергия, освобождаемая при движении иона натрия внутрь, обеспечивает движение другого вещества наружу. (Рис.6)
![]()
Котранспорт глюкозы и аминокислот с ионами натрия. Глюкоза и многие аминокислоты транспортируются внутрь большого числа клеток противзначительного градиента концентрации; осуществляется это исключительно механизмом котранспорта. Видно, что транспортный белок-переносчик имеет 2 участка связывания на его наружной стороне, 1 — для натрия и 1 — для глюкозы. Концентрация ионов натрия очень высокая снаружи клетки и очень низкая внутри, что обеспечивает энергию для транспорта. Транспортный белок обладает специфическим свойством: его кон формационное изменение не позволяет натрию двигаться внутрь клетки до тех пор, пока не присоединится молекула глюкозы. Когда прикрепляются оба вещества, автоматически происходит конформационное изменение белка-переносчика, в результате натрий и глюкоза одновременно транспортируются внутрь — это механизм котранспорта натрия и глюкозы. Котранспорт натрия и аминокислот происходит так же, как для глюкозы, но используются другие транспортные белки. Установлены 5 белков для транспорта аминокислоту каждый из которых способен транспортировать одну группу аминокислот со специфическими молекулярными характеристиками. Натриевый котранспорт глюкозы и аминокислот особенно характерен для эпителиальных клеток пищеварительного тракта и почечных канальцев и обеспечивает всасывание этих веществ в кровь, что будет обсуждаться в следующих главах. Другие важные механизмы котранспорта, обнаруженные в некоторых клетках, включают котранспорт ионов хлора, йода, железа и мочевой кислоты. Контртранспорт кальция и ионов водорода. Активный транспорт в тканях. Два особенно важных механизма контртранспорта (перенос в направлении, противоположном движению первичного иона) представлены натрий-кальциевым контртранспортом и натрий-водородным контртранспортом. Натрий-кальциевый контртранспорт осуществляется практически во всех клеточных мембранах, причем ионы натрия движутся внутрь, а ионы кальция — наружу, и оба иона связываются с одним транспортным белком, функционирующим в режиме контртранспорта. Этот механизм является дополнительным к первично активному транспорту кальция, который происходит в некоторых клетках. В ряде тканей осуществляется натрий-водородный контртранспорт. Наиболее важным примером являются проксимальные канальцы почек, где ионы натрия движутся из просвета канальца внутрь канальцевой клетки, тогда как протоны контртранспортом перемещаются в просвет канальца. В сравнении с первично активным транспортом протонов, происходящим в более дистальных почечных канальцах, роль контртранспортного механизма в концентрировании ионов водорода не столь значительна. Однако с его помощью может транспортироваться чрезвычайно большое число ионов водорода, что делает этот механизм ключевым для регуляции содержания этих ионов в жидкостях организма. (Рис.7)
![]()
Активный транспорт в тканях.Во многих частях организма вещества должны транспортироваться не просто через клеточную мембрану, а через слой клеток. Транспорт этого вида осуществляется через: (1) кишечный эпителий; (2) эпителий почечных канальцев; (3) эпителий экзокринных желез; (4) эпителий желчного пузыря; (5) мембрану хориоидного сплетения мозга и другие мембраны. Основными механизмами транспорта через клеточный слой являются: (1) активный транспорт через клеточную мембрану на одной стороне транспортирующих клеток; (2) простая или облегченная диффузия через мембрану на противоположной стороне этих клеток. На рисунке показан механизм транспорта ионов натрия через эпителиальный слой в кишечнике, желчном пузыре и почечных канальцах. На рисунке видно, что на стороне, обращенной в просвет, эпителиальные клетки плотно контактируют друг с другом посредством соединений, называемых легкими прикосновениями. Щеточная каемка на обращенной в просвет поверхности проницаема для ионов натрия и воды. В связи с этим натрий и вода легко диффундируют из просвета внутрь клетки. Затем ионы натрия активно транспортируются через базальную и латеральную клеточные мембраны во внеклеточную жидкость окружающей соединительной ткани и в кровеносные сосуды. В результате создается высокий градиент концентрации натрия через эти мембраны, что вызывает также осмос воды. Таким образом, активный перенос ионов натрия через базолатеральную мембрану эпителиальных клеток является причиной транспорта не только ионов натрия, но и воды. Эти механизмы обеспечивают поступление питательных веществ, ионов и других веществ из кишечника в кровь; таким же путем эти вещества реабсорбируются из клубочкового фильтрата в почечных канальцах.
Как происходит транспорт аминокислот в клетку?
Транспорт аминокислот через мембраны клеток , как при всасывании из полости кишечника в энтероциты, так и при переходе из крови в клетки различных тканей, осуществляется при помощи двух механизмов: вторичный активный транспорт и глутатионовая транспортная система.
Транспорт аминокислот через мембраны
Вторичный активный транспорт
Вторичный активный транспорт - это перенос веществ, в данном случае аминокислот, с использованием градиента концентрации натрия между внутренней и наружной сторонами клеточной мембраны.
Вторичный активный транспорт основан на наличии низкой концентрации ионов натрия внутри клеток, создаваемой мембранным ферментом Na + ,K + -АТФазой. Специфический белок-транспортер связывает на апикальной поверхности энтероцитов аминокислоту и ион натрия. Важно то, что в отсутствие натрия аминокислота не в состоянии связаться с белком-переносчиком.
Затем, изменив свое положение в мембране, белок отдает ион натрия в цитозоль по градиенту концентрации. Сразу после этого аминокислота теряет связь с белком и остается в цитоплазме.
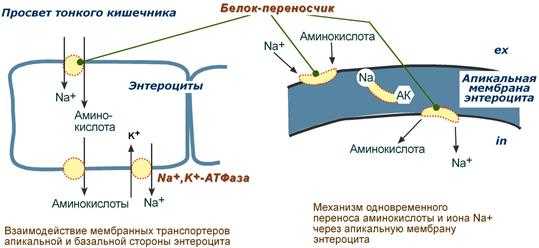
Вторичный активный транспорт аминокислот через мембраны
В настоящее время выделяют 5 транспортных систем:
- для крупных нейтральных, в том числе алифатических и ароматических аминокислот,
- для малых нейтральных - аланина, серина, треонина,
- для основных аминокислот - аргинина и лизина,
- для кислых аминокислот - аспартата и глутамата,
- для малых аминокислот - глицина, пролина и оксипролина.
Глутатионовая система транспорта
Второй способ переноса аминокислот внутрь клетки происходит в комплексе с глутатионом при помощи фермента γ-глутамилтрансферазы.
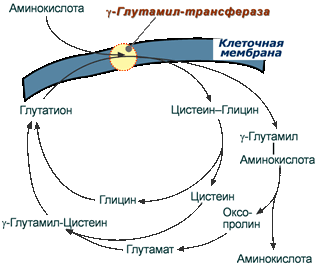
Транспорт аминокислот при участии глутатиона
Переносчиком некоторых аминокислот (обычно нейтральных) по этой схеме является трипептид глутатион (γ-глутамилцистеилглицин). При взаимодействии глутатиона с аминокислотой на внешней стороне клеточной мембраны γ-глутамильный остаток связывает аминокислоту и происходит ее перемещение внутрь клетки. Глутатион при этом распадается на составляющие. После отделения аминокислоты происходит ресинтез глутатиона.
Строение трипептида глутатиона
Исследование активности γ-глутамилтрансферазы часто проводится в клинико-лабораторной диагностике заболеваний печени и почек.
Читайте также:
- Фотодинамическая терапия (ФДТ), антагонисты СЭФР в терапии хориоидальной неоваскулярной мембраны (ХНВМ). Применение Луцентиса
- Результаты лечения гамма-ножом вестибулярной шванномы и прогноз
- Техника медиального доступа к пяточной кости
- Источники и факторы риска отравления свинцом
- Неиммунологические пути активации комплемента. Образование гранулем при аллергическом альвеолите