Взаимодействие вируса с клеткой. Характер взаимодействия вирус-клетка. Продуктивное взаимодействие. Вирогения. Интерференция вирусов.
Добавил пользователь Дмитрий К. Обновлено: 28.01.2026
Взаимодействие идет в единой биологической системе на генетическом уровне.
Существует четыре типа взаимодействия:
1) продуктивная вирусная инфекция (взаимодействие, в результате которого происходит репродукция вируса, а клетки погибают);
2) абортивная вирусная инфекция (взаимодействие, при котором репродукции вируса не происходит, а клетка восстанавливает нарушенную функцию);
3) латентная вирусная инфекция (идет репродукция вируса, а клетка сохраняет свою функциональную активность);
4) вирус-индуцированная трансформация (взаимодействие, при котором клетка, инфицированная вирусом, приобретает новые, ранее не присущие ей свойства).
После адсорбции вирионы проникают внутрь путем эндоцитоза (виропексиса) или в результате слияния вирусной и клеточной мембран. Образующиеся вакуоли, содержащие целые вирионы или их внутренние компоненты, попадают в лизосомы, в которых осуществляется депротеинизация, т. е. «раздевание» вируса, в результате чего вирусные белки разрушаются. Освобожденные от белков нуклеиновые кислоты вирусов проникают по клеточным каналам в ядро клетки или остаются в цитоплазме.
Нуклеиновые кислоты вирусов реализуют генетическую программу по созданию вирусного потомства и определяют наследственные свойства вирусов. С помощью специальных ферментов (полимераз) снимаются копии с родительской нуклеиновой кислоты (происходит репликация), а также синтезируются информационные РНК, которые соединяются с рибосомами и осуществляют синтез дочерних вирусных белков (трансляцию).
После того как в зараженной клетке накопится достаточное количество компонентов вируса, начинается сборка вирионов потомства. Процесс этот происходит обычно вблизи клеточных мембран, которые иногда принимают в нем непосредственное участие. В составе вновь образованных вирионов часто обнаруживаются вещества, характерные для клетки, в которой размножается вирус. В таких случаях заключительный этап формирования вирионов представляет собой обволакивание их слоем клеточной мембраны.
Последним этапом взаимодействия вирусов с клетками является выход или освобождение из клетки дочерних вирусных частиц. Простые вирусы, лишенные суперкапсида, вызывают деструкцию клетки и попадают в межклеточное пространство. Другие вирусы, имеющие липопротеидную оболочку, выходят из клетки путем почкования. При этом клетка длительное время сохраняет жизнеспособность. В отдельных случаях вирусы накапливаются в цитоплазме или ядре зараженных клеток, образуя кристаллоподобные скопления - тельца включений.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
ВЗАИМОДЕЙСТВИЕ МУРАВЬЕВ В СЕМЬЕ
ВЗАИМОДЕЙСТВИЕ МУРАВЬЕВ В СЕМЬЕ Семья муравьев — объединение сотен, тысяч, а иногда и миллионов индивидов, усилиями которых сооружается гнездо, выкармливается многочисленное потомство, охраняется муравейник и его кормовой участок от посягательств агрессивных
Что это за плазматические клетки, вырабатывающие антитела, и можно ли плазматическую клетку считать самой главной клеткой иммунной системы?
Что это за плазматические клетки, вырабатывающие антитела, и можно ли плазматическую клетку считать самой главной клеткой иммунной системы? — Что это за плазматические клетки, вырабатывающие антитела? О них уже знали во времена Мечникова или это более позднее
10. Морфология вирусов, типы взаимодействия вируса с клеткой
10. Морфология вирусов, типы взаимодействия вируса с клеткой Вирусы - микроорганизмы, составляющие царство Vira.Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).По форме вирионы могут быть: округлыми, палочковидными, в виде
Взаимодействие наследственности и среды
Взаимодействие наследственности и среды Иногда спрашивают: что важнее — наследственность или окружающая среда? На этот вопрос не так легко ответить. Если под этим подразумевать, чт? имеет наибольшую силу воздействия, то и тогда следует ограничиться частными случаями.
ВОЗВРАЩЕНИЕ ХОЗЯИНА
ВОЗВРАЩЕНИЕ ХОЗЯИНА Приветливый хозяин, ведущий Чосера и других странников из Лондона в Кентербери, и бывший посредником в их рассказах, повернул назад и привел их обратно к Лондону. Если я теперь возвращаюсь к современности, то исключительно потому, что следование одним
Прощание Хозяина
Прощание Хозяина Если по возвращении хозяина я размышляю обо всем путешествии, благодарным участником которого я был, моей непреодолимой реакцией является что-то вроде изумления. Изумление не только от феерии деталей, которые мы увидели; изумление также от самого факта,
7. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ
7. ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровне. Особенно широка интеграция сигналов в ретикулярной формации. В коре мозга происходит интеграция сигналов высшего порядка. В
4.3. Взаимодействие генов
4.3. Взаимодействие генов В организме одновременно функционирует множество генов. В процессах реализации генетической информации в признак возможны многочисленные «пункты» взаимодействия разных генов на уровне биохимических реакций. Такие взаимодействия неизбежно
4.4. Взаимодействие генотипа и среды
4.4. Взаимодействие генотипа и среды Природа проявления действия генов намного сложнее, чем в описанных выше вариантах. Рассматривая действие генов и их аллелей, необходимо учитывать влияние внешней среды на проявление признаков, а также модифицирующее действие других
Взаимодействие локусов А и Е
Взаимодействие локусов А и Е Важность этих двух серий аллелей будет показана в данном разделе. Обе серии контролируют распределение черного и желтого пигментов по шерсти у большинства пород собак. Во взаимодействии эти локусы образуют множество хорошо известных
Взаимодействие и интеграция
Взаимодействие и интеграция В наши дни посетители музеев воспринимают выставленные в них смонтированные скелеты огромных вымерших животных как нечто само собой разумеющееся. Однако так было не всегда. Научные сведения о таких импозантных формах, как мамонты и
2.2.4. Определение способа взаимодействия хозяина с собакой
2.2.4. Определение способа взаимодействия хозяина с собакой Поскольку зачастую установлением причины девиантного поведения животного занимается врач-ветеринар общего профиля, который не обладает профессиональной психологической подготовкой, автором предлагается тест,
9.7. Заключительные замечания (взаимодействие в биоценозах)
9.7. Заключительные замечания (взаимодействие в биоценозах) Рассмотрим биохимические взаимодействия живых организмов в естественных условиях, которые распространены чрезвычайно широко и стали объектом специальной науки — аллелопатии (см. гл. 1). Примеры таких
Возвращение Хозяина Трактирщик, сопровождавший Чосера и других пилигримов в Кентербери и исполнивший роль импресарио, привел их обратно в Лондон. И если я теперь вернусь в настоящее, оно должно быть иным: ожидание того, что эволюция дважды проследует одним путем,
Прощание Хозяина Я восхищаюсь паломничеством, в котором сам с благодарностью участвовал. Восхищение у меня вызывает не только торжество жизни, но и то обстоятельство, что на некоей планете вообще может существовать жизнь. Вселенная могла бы остаться безжизненной, с
Типы взаимодействия вируса с клеткой. Фазы репродукции вирусов
Типы взаимодействия вируса с клеткой.Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Репродукция вирусов осуществляется в несколько стадий, последовательно сменяющих друг друга: адсорбция вируса на клетке; проникновение вируса в клетку; «раздевание» вируса; биосинтез вирусных компонентов в клетке; формирование вирусов; выход вирусов из клетки.
Адсорбция.Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку.Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
«Раздевание».Процесс «раздевания» заключается в удалении защитных вирусных оболочек и освобождении внутреннего компонента вируса, способного вызвать инфекционный процесс. «Раздевание» вирусов происходит постепенно, в несколько этапов, в определенных участках цитоплазмы или ядра клетки, для чего клетка использует набор специальных ферментов. В случае проникновения вируса путем слияния вирусной оболочки с клеточной мембраной процесс проникновения вируса в клетку сочетается с первым этапом его «раздевания». Конечными продуктами «раздевания» являются сердцевина, нуклеокапсид или нуклеиновая кислота вируса.
Биосинтез компонентов вируса.Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Формирование (сборка) вирусов.Синтезированные вирусные нуклеиновые кислоты и белки обладают способностью специфически «узнавать» друг друга и при достаточной их концентрации самопроизвольно соединяются в результате гидрофобных, солевых и водородных связей.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Выход вирусов из клетки.Различают два основных типа выхода вирусного потомства из клетки. Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки. Второй тип — почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции.
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
39. Типы взаимодействия вируса с клеткой. Стадии репродукции вирусов.
Типы взаимодействия вируса с клеткой. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
Продуктивный тип— завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип— не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения— характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Репродукция вирусовосуществляется в несколько стадий, последовательно сменяющих друг друга: адсорбция вируса на клетке; проникновение вируса в клетку; «раздевание» вируса; биосинтез вирусных компонентов в клетке; формирование вирусов; выход вирусов из клетки.
Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
«Раздевание». Процесс «раздевания» заключается в удалении защитных вирусных оболочек и освобождении внутреннего компонента вируса, способного вызвать инфекционный процесс. «Раздевание» вирусов происходит постепенно, в несколько этапов, в определенных участках цитоплазмы или ядра клетки, для чего клетка использует набор специальных ферментов. В случае проникновения вируса путем слияния вирусной оболочки с клеточной мембраной процесс проникновения вируса в клетку сочетается с первым этапом его «раздевания». Конечными продуктами «раздевания» являются сердцевина, нуклеокапсид или нуклеиновая кислота вируса.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Формирование (сборка) вирусов. Синтезированные вирусные нуклеиновые кислоты и белки обладают способностью специфически «узнавать» друг друга и при достаточной их концентрации самопроизвольно соединяются в результате гидрофобных, солевых и водородных связей.
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
Выход вирусов из клетки. Различают два основных типа выхода вирусного потомства из клетки. Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки. Второй тип — почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции.
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции. Продуктивная вирусная инфекцияс образованием дочерних популяций и характерными клиническими проявлениями возможна лишь при наличии в заражённом организме чувствительных клеток, в которых осуществляется репродуктивный цикл возбудителя. Например, возбудитель полиомиелита может реплицировать только в клетках ЖКТ и ЦНС приматов и человека.
Абортивная инфекция развивается при проникновении возбудителя в нечувствительные клетки (например, при попадании вируса лейкоза коров в организм человека) либо в клетки, не способные обеспечить полный репродуктивный цикл (например, находящиеся в стадии клеточного цикла G0). Способность клеток к поддержанию вирусспецифических репродуктивных процессов также подавляет ИФН, противовирусный эффект которого направлен против самых различных вирусов.
Персистирующая вирусная инфекция возникает при таком взаимодействии между вирусом и заражённой клеткой, когда в последней продолжается выполнение собственных клеточных функций. Если заражённые клетки делятся, образуется инфицированный клон. Таким образом, увеличение числа заражённых клеток способствует увеличению общей популяции возбудителя в организме. Тем не менее персистирующие вирусные инфекции обычно нарушают функции клеток, что в конце концов приводит к клиническим проявлениям. У человека развитие персисти-рующих инфекций в определённой степени зависит от возраста. Например, внутриутробное заражение вирусом коревой краснухи или цитомегаловирусом (ЦМВ) приводит к ограниченному по времени персистированию возбудителя. Появление симптоматики связано с возможностью плода развивать иммунные реакции на инфекционный агент.
Латентная (скрытая) вирусная инфекция. В то время как персистирующие инфекции сопровождаются постоянным высвобождением дочерних вирусных популяций, при латентных поражениях они образуются спорадически. Репродуктивный цикл подобных возбудителей резко замедляется на поздних стадиях и активируется под влиянием различных факторов. Латентные инфекции характерны для большинства герпесвирусов, вызывающих рецидивирующие и обычно не прогрессирующие заболевания.
Инаппарантные инфекции [от лат. in-, отрицание, + арраrео, являться] сопровождаются бессимптомной циркуляцией незначительных количеств возбудителя в отдельных органах. При этом выявить возбудителя можно лишь специальными методами. От бессимптомного носительства подобные поражения отличает большая вероятность возникновения клинических проявлений. Этот термин применяют при целом ряде инфекций, при которых нет явных признаков заболевания. В практике вирусных инфекций у человека часто применяют альтернативный термин «субклиническая инфекция». Собственно, и латентные инфекции можно расценивать как хронически протекающие инаппаратные инфекции, при которых устанавливается баланс между организмом и возбудителем.
Дремлющая (криптогенная) вирусная инфекция — форма проявления вирусной инфекции при которой возбудитель в неактивном состоянии находится в отдельных очагах (например, в нервных ганглиях). Клинически инфекция проявляется лишь при резком ослаблении защитных сил организма. Например, вирус герпеса 3 типа, вызывающий при первичном заражении ветряную оспу, пожизненно сохраняется в организме. Рецидив заболевания в форме опоясывающего лишая возможен лишь при нарушениях иммунного статуса (наиболее часто в пожилом возрасте).
Медленные вирусная инфекции характеризуются длительным инкубационным периодом (месяцы и годы), в течение которого возбудитель размножается, вызывая всё более явные повреждения тканей. Первоначально возбудитель размножается в ограниченной группе клеток, но постепенно инфицирует всё большее их число. Заболевания заканчиваются развитием тяжёлых поражений и смертью больного. К медленным вирусным инфекциям относят подострый склерозирующий панэнцефалит, ВИЧ-инфекцию и др.
ВЗАИМОДЕЙСТВИЕ ВИРУСОВ С КЛЕТКАМИ
Вирусные частицы, или вирионы являются инертными, статическими формами вируса. Когда вирионы находятся вне клетки, они не репродуцируются. В настоящее время известно три типа взаимодействия вирусов с клетками хозяина.
Продуктивный тип взаимодействия заканчивается образованием вирусного потомства.
Абортивный тип не завершается образованием вирусных частиц, поскольку инфекционный процесс прерывается и не сопровождается формированием инфекционного потомства.
Интегративный тип взаимодействия вирусов с клеткой характерен для онкогенных вирусов, нуклеиновая кислота которых способна встраиваться (интегрироваться) в клеточную хромосому, вызывая трансформацию клеток. Границы между вирусами с автономной репликацией геномов и интеграционными вирусами довольно условны, и один и тот же вирус, в зависимости от вида клеток, может вести себя либо как инфекционный, либо как интеграционный геном. Результатом такого взаимодействия вируса и клетки является изменение наследственных свойств клетки. Данный тип взаимодействия вируса и клетки называют вироге-
нией, подобно лизогении при взаимодействии фагов с бактериями. Вирусы, способные вызывать вирогению, относят к группе умеренных.
Продуктивный тип взаимодействия вируса и клетки получил название репродукции вирусов (от англ. rерroduсе — воспроизводить). Репродукция вирусов — это образование по принципу комплементарности и путем репликации копий вирусных нуклеиновых кислот и индуцирование молекулами последних биосинтеза вирусных белков с последующей самоорганизацией этих компонентов в вирусные частицы.
Синтез и репликацию нуклеиновых кислот вирусов осуществляют ферменты. Используя нуклеотиды клетки, ферменты создают из них полинуклеотидные цепи новых молекул нуклеиновых. кислот вирусов. В зависимости от типа синтезируемых нуклеиновых кислот они называются ДНК-полимеразами или РНК-синтетазами. В некоторых случаях нуклеиновые кислоты вирусов реплицируются клеточными полимеразами — ферментами, присутствующими в клетке до ее заражения вирусом, хотя чаще репликацию вирусных нуклеиновых кислот осуществляют полимеразы, появляющиеся после заражения клетки вирусом. Такие полимеразы называют вирусспецифическими, так как биосинтез их закодирован в структуре нуклеиновых кислот самих вирусов. И, наконец, репликация нуклеиновых кислот некоторых вирусов происходит за счет предсуществующих в вирионе полимераз. Синтез вирусспецифических полимераз и вирусных структурных белков осуществляется на рибосомах клетки. Вирусные полимеразы, например РНК-репликазы, являются строго специфичными.
При большом разнообразии механизмов репродукции вирусов общим для них является то, что источником мономеров для синтеза и репликации нуклеиновых кислот служат нуклеотиды клетки. Источником мономеров для синтеза и построения белков всех вирусов являются аминокислоты, и синтез белков всех вирусов независимо от ультраструктуры их нуклеиновых кислот осуществляется в клеточных рибосомах. Источником энергии для биосинтетических процессов при репродукции вирусов является АТФ, которая вырабатывается в митохондриях клетки-хозяина.
Процесс репродукции вирусов включает шесть этапов: адсорбцию вируса на клетке; проникновение в клетку; депротеинизацию и освобождение вирусного генома; синтез вирусных компонентов в клетке-хозяине; сборку и формирование вирусов; выход зрелых вирусов из клетки.
Адсорбция, то есть прикрепление ви руса к клетке, осуществляется специфическими и неспецифическими механизмами. Неспецифический — определяется силами электростатического
взаимодействия. В этом процессе участвуют положительно заряженные аминные группы вирусного белка и кислые фосфатные группы клеточной поверхности, имеющие отрицательный заряд. Специфический механизм взаимодействия вируса и клетки обусловлен комплементарными клеточными и вирусными рецепторами. Вирусные рецепторы подразделяют на липопротеиновые (у арбовирусов) и мукопротеиновые (у миксовирусов и аденовирусов). Спектр чувствительности клеток к вирусам часто определяется наличием соответствующих рецепторов.
Резистентность клеток можно преодолеть путем разрушения клеточной мембраны, для чего используют инактивированный вирус Сендай. Специфические противовирусные антитела и антитела к нормальным клеткам препятствуют адсорбции вирусов. Процесс адсорбции состоит из двух периодов — обратимого и необратимого. Период обратимой адсорбции может закончиться десорбцией вируса. При длительном контакте клеток и вируса наступает стадия необратимой адсорбции. Адсорбированные вирионы таким образом могут частично элюироваться с поверхности клеток, небольшая часть остается интактной, а основная масса вирусных частиц проникает в клетку.
Проникновение вирусов в клетку осуществляется путем ви-ропексиса, или пиноцитоза. При этом в месте адсорбции вириона происходит сначала инвагинация наружной мембраны клетки и образование внутриклеточной вакуоли с вирусной частицей. Через некоторое время вирусная и клеточная мембраны вакуоли лизируются и высвобождается нуклеокапсид вируса. Другие вирусы, например вирус Неrреs simр1ех, проникают в клетку не путем виропексиса, а расплавления мембран вируса и клетки. После расплавления вирусный нуклеопротеид оказывается в цитоплазме. Большинство вирусов проникают в клетку путем виропексиса, меньшинство видов — путем сплавления. У фагов этот процесс, то есть освобождение ДНК и ее последующая инъекция сквозь оболочку бактериальной клетки, происходит непосредственно на поверхности этой клетки сразу же после прикрепления к ней фага.
Депротеинизация, или «раздевание» происходит постепенно в несколько этапов. Процесс освобождения вирусной нуклеиновой кислоты идет при активном участии самого вируса, индуцирующего образование в клетке ферментов и активаторов ферментов, необходимых для депротеинизации вирусной нуклеиновой кислоты. В процессе депротеинизации вирусов могут участвовать ферменты клеточных лизосом. Таким образом, депротеинизация завершается освобождением вирусного генома.
Как только вирусный геном освобождается от белка, вирусная нуклеиновая кислота дезорганизует работу клеточных систем. Вирус стимулирует синтез ингибитора клеточных РНК. Ингиби-
тор представляет собой белок — гистон, кроме того блокирующий процесс синтеза клеточной ДНК. Второй «ранний» вирусный белок препятствует осуществлению функции клеточной и-РНК. Термин «ранние» обозначает молекулы, синтезированные до репликации ДНК.
Реализация генетической информации вирусов осуществляется в соответствии с процессами транскрипции — синтеза информационных РНК, комплементарных матричным ДНК или РНК; трансляции — синтеза белков на рибосомах клетки с участием и-РНК; репликации — синтеза молекул нуклеиновой кислоты, гомологичных геному (рис. 7).
| Синтез вирусных ДНК у ДНК-содержащих вирусов осуществляется с помощью ДНК-полимераз. Благодаря участию этого фермента из нуклеотидов клетки синтезируется и строится вторая комплементарная нить ДНК, в результате чего образуются новые двухцепочечные молекулы ДНК. Процесс репликации молекул ДНК продолжается до тех пор, пока в клетке не накопится определенное их количество, необходимое для потомства вирусной частицы, проникшей в клетку.
Поскольку типы и формы нуклеиновых кислот разнообразны — кроме двухцепочечной ДНК могут быть РНК, одно- и двухцепочечные, линейные и кольцевые молекулы — то и механизмы их репликации различны.
У вирусов, содержащих двунитевуюРНК, синтез вирусных компонентов происходит сходным образом. «Ранняя» транскрипция у них осуществляется благодаря вирионному ферменту — РНК-зависимой РНК-полимеразе. Вирусы с однонитевой РНК по характеру синтеза белков разделяются на две группы: вирусы,
| Вирусные белки в т. ч. ферменты |
| ОАО |
| Новые молекулы вирусной ДНК |
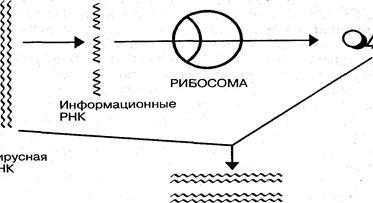
Рис. 7. Схема репликациодвухцепочечных молекул вирусных ДНК и передачи генетической информации в рибосомы
| И-РНК(+) |
| Вирусные белки в т. ч. РНК-репликаза |
| Транскриптаза ———————^ Вирусная РНК |
 |
| ОАО |
| Новые молекулы вирусных(-) РНК |
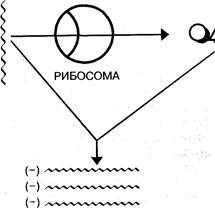
Рис. 8. Схема репликации вирусных минус-РНК и передачи генетической информации в рибосомы
у которых РНК обладает информационными свойствами и направляет синтез специфических белков (пикорна- и тогавирусы);
вирусы, у которыхРНК не является информационной, а служит только матрицей для синтезаи-РНК (ортомиксо-, парамиксо-, рабдовирусы) (рис. 8).
Особым способом репликации отличаются РНК-содержащие двухцепочечные ретровирусы. Существенное отличие состоит в том, что у них имеется фермент — РНК-зависимая-ДНК-полимераза, с помощью которой на матрице вирионной РНК синтезируется ДНК-провирус кольцевой формы. Этот провирус встраивается в клеточную хромосому и транскрибируется клеточной РНК-полимеразой точно так же, как и обычные клеточные гены. Образующаяся РНК направляет синтез вирусных белков и одновременно является вирионной РНК. Вирусная нуклеиновая кислота кодирует синтез двух классов белков: неструктурных белков — ферментов, которые обеспечивают процесс репродукции вирусов на разных его этапах, и структурных белков, которые войдут в состав вирионов потомства. Наряду с синтезом белков в клетке при репродукции вирусов, имеющих суперкапсид, например, вируса гриппа, происходит синтез углеводных цепей, входящих в состав гликопротеидов. Присоединение углеводов осуществляется с помощью клеточных ферментов трансфераз, синтез липидов суперкапсида — клеткой-хозяином. Вирусный суперкапсид формируется при включении липидов из плазматической мембраны клетки-хозяина. Синтез вирусных нуклеиновых кислот и вирусспецифических белков происходит почти одновре-
менно и не менее чем на час опережает созревание вирусных частиц.
Сборка и формирование вирионов осуществляется после достижения критической концентрации нуклеиновой кислоты и белка так как чисто физико-химическая реакция агрегации белка с вирусными нуклеиновыми кислотами. Так, у парамиксовирусов формирование вирионов происходит путем самосборки, которая обусловлена «узнаванием» РНК белками. Вероятно, «узнающим» белком вляется белок Р, который наиболее жестко связан с РНК в вирионах. Нуклеокапсиды накапливаются в цитоплазме зараженных леток, причем скорость образования внутриклеточных нуклеокапсидов гораздо выше скорости образования вируса. Зрелые вирусные сложноустроенные частицы формируются при прохождении их нуклеопротеидов через цитоплазматическую или ядерную мембрану клетки-хозяина.
Компоненты этих мембран становятся элементами наружной оболочки вириона — суперкапсида. Заключительная стадия репро-дукции вирусов — выход вновь сформированных вирионов из клетки. У различных групп вирусов этот процесс происходит по-разному. При выходе из клетки вирусов, не имеющих суперкапсидной оболочки, как правило, клетка погибает из-за взрывного выхода одновременно большого количества вирусных частиц. Вирусы, имеющие суперкапсидную оболочку, выходят из клетки путем почкования. На заключительном этапе сборки нуклеокапсиды сложноустроенных вирусов фиксируются на клеточной мембране, модифицированной вирусными белками, и постепенно выпячивают ее, в результате чего образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки, то есть внешняя суперкапсидная оболочка этих вирусов формируется в процессе их выхода из клетки.
При таком механизмевыхода вирионов из клетки последняя сохраняет в определенной мере свои функции. Так, например, процесс выхода вирусов гриппа может продолжаться более 30 часов. Герпесвирусы могут выходить из клетки через цитоплазматические трубочки, соединяющие ядерную оболочку с наружной мембраной клетки. Благодаря такому механизму выделения эти вирусы передаются от клетки к клетке, не выходя во внешнюю среду. Образовавшиеся в процессе репродукции вирионы могут инфицировать новые клетки и проходить в них новый цикл репродукции.
Интегративный тип взаимодействия характеризуется встраиванием нуклеиновой кислоты вируса в хромосому клетки-хозяина. При этом вирусный геном функционирует как составная часть клеточного генома. Интегративный тип взаимодействия присутствует у бактериофагов, онкогенных вирусов, вируса гепатита В, вирусов герпеса и вируса иммунодефицита человека. В частности,
| 5 «Микробиология» |
умеренные фаги вступают в симбиоз с частью бактерий, при этом ДНК фага встраивается в хромосому бактерии. В данном случае геномом фага называют профаг, ставший частью хромосомы бактерии и не вызывающий ее лизиса.
Симбиоз микробной клетки с умеренным фагом называют лизогенией. Самопроизвольно или под действием ультрафиолета, или химических факторов профаг может из хромосомы переходить в цитоплазму и вести себя как вирулентный фаг, лизирующий бактерии.
У онкогенных вирусов и вируса иммунодефицита человека процесс интеграции является обязательным в цикле их репродукции. У этих вирусов на матрице РНК с помощью фермента РНК-зависимой ДНК-полимеразы синтезируется ДНК-копия, которая встраивается в хромосому клетки. ДНК вируса, находящаяся в составе хромосомы клетки, называется ДНК-провирусом. Клетка сохраняет свои функции и при делении передает ДНК-провирус дочерним клеткам.
Таким образом, состояние вирогении наследуется. ДНК-про-вирус несет дополнительную генетическую информацию, поэтому интеграция является причиной некоторых аутоиммунных и хронических заболеваний, опухолей. Под действием некоторых физических и химических факторов ДНК-провирус может вырезаться из клеточной хромосомы и переходить в автономное состояние, включаясь в цикл репродукции.
Тема 10. Основы вирусологии. Бактериофагия.
Вирусы бактерий: структура, распространение, практическое применение в медицине.
МОРФОЛОГИЯ И СТРУКТУРА ВИРИОНОВ
Предметом изучения медицинской вирусологии является: вирусы - паразиты человека, их морфология, физиология, генетика, экология и др.
Вирусы - неклеточные формы жизни, имеющие геном, окруженный белковой оболочкой, являющиеся облигатными паразитами. В настоящее время известны вирусы бактерий, грибов, растений, животных.
Внеклеточная форма - вирион - включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.).
Внутриклеточная форма - вирус - может быть представлена лишь одной молекулой нуклеиновой кислоты.
По морфологии выделяют вирусы палочковидные, пулевидные, сферические, овальные, комбинированные.
По размерам вирусы бывают от крупных (до 400 нм) до мелких (20-30 нм).
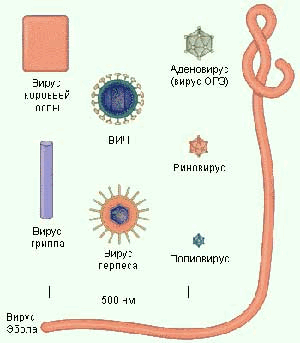
Рис. 1. Морфология вирусов.
В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов - сложные вирусы (одетые), либо в его состав входят только нуклеиновые кислоты и белки - простые вирусы (голые).
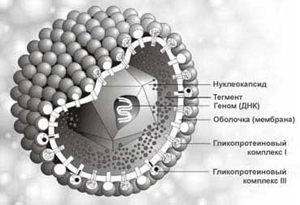
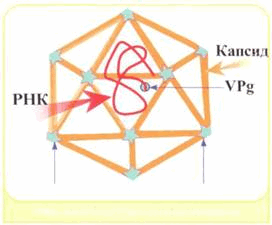
Рис. 2. Структура сложного вируса (слева); структура простого вируса (справа), белковая оболочка - капсид, внутри находится нуклеиновая кислота; стрелками снизу указаны вершины пентамера.
Вирусы содержат только один тип нуклеиновой кислоты, ДНК или РНК, но не оба типа одновременно.
Капсид - белковая капсула защищающая геном. Капсид образуют одинаковые по строению субъединицы -капсомеры, организованные в один или два слоя по двум типам симметрии - кубическому или спиральному .
Организация по принципу спиральной симметрии придает вирусам палочковидную форму.
Организация по принципу кубической симметрии придает вирусам сферическую форму.
Нуклеокапсид - комплекс капсида и вирусного генома. В состав нуклеокапсидов также входят внутренние белки, обеспечивающие правильную упаковку генома, а также выполняющие структурную и ферментативную функции.
Суперкапсид - особая оболочка, организованная двойным слоем липидов и вирусными белками, покрывающая капсид (имеют сложные вирусы).
Рис. 3. Капсид вируса табачной мозаики построен по спиральному типу симметрии: электонная микрофотография (слева), структура нуклеокапсида (справа).
Рис. 4. Капсид вируса гепатита А (слева) и аденовируса (справа) построены по кубическому типу симметрии.
КЛАССИФИКАЦИЯ ВИРУСОВ
Вирусы отнесены к царству Vira. В основу классификации положен тип нуклеиновой кислоты. Выделяют рибовирусы (РНК-содержащие вирусы) и дезоксирибовирусы (ДНК-содержащие вирусы). Для вирусов разработаны следующие таксономические категории (по восходящей): Вид - Род - Подсемейство - Семейство - Подцарство - Царство.
При систематизировании вирусов выделяют следующие основные критерии: сходство нуклеиновых кислот, размеры, наличие или отсутствие суперкапсида, тип симметрии нуклеокапсида, характеристика нуклеиновой кислоты и так далее.
ВЗАИМОДЕЙСТВИЕ ВИРУСА С КЛЕТКОЙ ХОЗЯИНА
(особенности вирусных инфекций)
Вирусы не способны самостоятельно размножаться. Синтез вирусных белков и воспроизведение копий вирусного генома обеспечивают биосинтетические процессы клетки-хозяина. При этом белковые макромолекулы и нуклеиновые кислоты образуются отдельно, после чего происходит самосборка дочерних популяций. Такой тип репродукции называется дизъюнктивный (разобщенный).
Реализация репродуктивного цикла в существенной степени зависит от типа инфицирования клетки и характера взаимодействия вируса с чувствительной (могущей быть инфицированной) клеткой.
Известны следующие типы взаимодействий «вирус-клетка».
Продуктивный (образуется дочерняя популяция).
Абортивный (дочерняя популяция не образуется).
Продуктивное взаимодействие «вирус-клетка» чаще носит литический характер, то есть заканчивается гибелью инфицированной клетки, что происходит после полной сборки дочерней популяции и выхода вирусов из клетки.
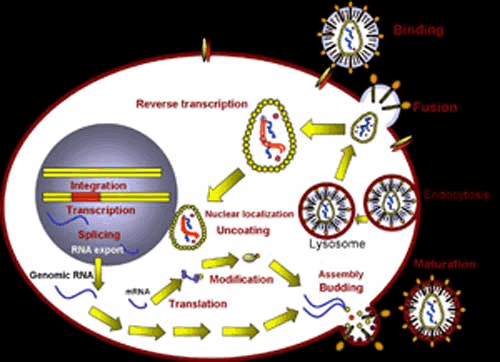
Рис. 5. Репродукция ВИЧ. Вирус прикрепляется к рецепторам клетки и, освободившись от суперкапсида, проникает в клетку. РНК вируса, проникнув в ядро, встраивается в ДНК после того, как с помощью фермента обратной транскриптазы РНК вируса трансформируется в ДНК. В дальнейшем на рибосомах синтезируются вирусные белки, образующие капсид. Происходит транскрипция РНК. Формируются вирионы, которые "одеваются" в мембрану клетки, выходя из нее.
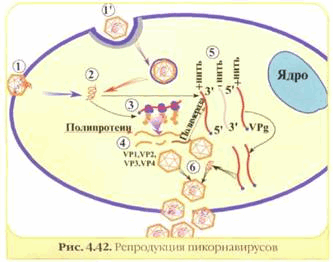
Рис. 6. Репродукция простого вируса.
Интегративное взаимодействие, или вирогения не приводит к гибели клетки. Нуклеиновая кислота вируса встраивается (интегрирует) в геном клетки-хозяина и в последующем функционирует как его составная часть.
Вирус, интегрированный в клеточный геном, называется провирус . При этом вирусная частица может стать неактивной, иногда остается в клетке очень долго, ничем не выдавая своего присутствия. Такие бессимптомные, скрытые инфекции называются латентными. Неблагоприятные для организма воздействия холодом, рентгеновскими лучами, химическими веществами и т.п. может возбудить их агрессивность. Возможны периодические реактивации с переходом в продуктивное взаимодействие «вирус-клетка», либо клетка трансформируется, давая начало злокачественному росту (онкогенные вирусы) .
Прионы - это белковые инфекционные агенты, кодирующиеся в нормальной или мутантной форме геномом хозяина, не обладающие автономными механизмами репликации и вызывающие в процессе развития инфекционного процесса накопление фибриллярных белков, ассоциирующихся в амилоид. Прионы - возбудители летальных дегенеративных изменений центральной нервной системы человека и животных. Прионы высокоустойчивы к действию ионизирующих и УФ-лучей, ультразвука, к высоким температурам и многим дезинфектантам. Разрушаются под действием эфира и фенола. Прионы могут персистировать в организме хозяина длительное время (годами и десятилетиями) не вызывая иммунного ответа.
Прионные инфекции - категория трансмиссивных нейродегенеративных болезней животных и человека из группы медленных вирусных инфекций.
ВИРУСЫ БАКТЕРИЙ (БАКТЕРИОФАГИ)
Бактериофаги (фаги) - вирусы, обладающие способностью проникать в бактериальные клетки, репродуцироваться в них и вызывать их лизис. Большинство фагов имеют сперматозоидную форму. Они состоят из головки, которая содержит нуклеиновую кислоту и отростка. Большинство из фагов содержат двунитевую ДНК, которая замкнута в кольцо.
Проникновение фага в бактериальную клетку происходит путем инъекции нуклеиновой кислоты через канал отростка.
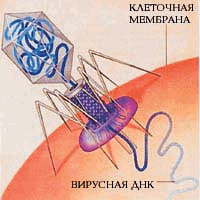
Рис. 7. Бактериофаги на бактерии (слева); бактериофаг вводит нуклеиновую кислоту в клетку бактерии (справа).
Выход зрелых фагов из бактериальной клетки происходит путем «взрыва», зараженные бактерии лизируются (вирулентные фаги). Лизогения - тип взаимодействия фага с бактериальной клеткой, при котором бактериофаг встраивает свою ДНК в бактериальный геном. Фаги, вызывающие данный тип инфекции, получили название умеренных. Фаговая ДНК, ассоциированная с геномом своего хозяина, носит название профаг .
Фаги широко распространены в природе. Фаги более устойчивы к действию физических и химических факторов, чем многие вирусы человека.
Практическое применение фагов в медицине. Препараты бактериофагов выпускают в жидком виде, в таблетках, в форме мазей, аэрозолей и свечей и применяют для лечения и профилактики.
Строгая специфичность бактериофагов позволяет использовать их для фаготипирования и дифференцировки бактериальных культур (фагодиагностика).
КУЛЬТИВИРОВАНИЕ ВИРУСОВ
В вирусологической практике для репродукции вирусов с диагностическими целями используются организмы чувствительных животных, куриные эмбрионы и культуры клеток, полученные из нормальных и злокачественных клеток людей и животных.
Культивирование вирусов помогает решить ряд теоретически проблем, связанных с изучением особенностей взаимодействия "вирус-клетка". Кроме того, решение целого ряда прикладных задач, связанных с диагностикой и производством препаратов для профилактики вирусных инфекций невозможно без накопления вируссодержащего сырья.
Читайте также:
- Вещества с бензолоподобным действием на кровь. Хлорбензол и гексахлорциклогексан
- Фертильность после хирургического лечения внематочной беременности. Рецидив
- Кровоизлияние, катаракта после операции на диабетической отслойке сетчатки. Тактика
- Колоноскопия при высоком риске инфекционных осложнений. Принципы
- Механизм транспорта COPII окаймленными везикулами