
Лабораторная работа вирусные заболевания у растений
Существует большая группа живых существ, не имеющих клеточного строения. Эти существа носят названия вирусов (от лат. "вирус" — яд) и представляют неклеточные формы жизни. Вирусы нельзя отнести ни к животным, ни к растениям. Они исключительно малы, поэтому могут быть изучены только с помощью электронного микроскопа.
Вирусы способны жить и развиваться только в клетках других организмов. Вне клеток живых организмов вирусы жить не могут, и многие из них во внешней среде имеют форму кристаллов
ВВЕДЕНИЕ 3
1. ИСТОРИЯ ОТКРЫТИЯ ВИРУСОВ 4
2. ПРИРОДА И ПРОИСХОЖДЕНИЕ ВИРУСОВ 5
2.1. Размеры вирусов 6
2.2. Поведение 6
3. ВИРУСЫ КАК ВОЗБУДИТЕЛЬ ЗАБОЛЕВАНИЙ У РАСТЕНИЙ 7
3.1. Распространение вирусов по растению 7
3.2. Механизмы передачи вирусов растений 10
3.3. Передача вирусов растений членистоногими переносчиками 11
3.4. Передача вирусов растений нематодами и грибами 13
4. ОСНОВНЫЕ ВИДЫ ВИРУСНЫХ ЗАБОЛЕВАНИЙ РАСТЕНИЙ 15
4.1. Мозаичность листьев 15
4.2. Курчавость листьев 15
4.3. Пожелтение листьев 16
5. МЕРЫ БОРЬБЫ С ВИРУСНЫМИ ЗАБОЛЕВАНИЯМИ 22
ВЫВОДЫ 23
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ 25
курсовая по биологии.doc
Министерство образования и науки, молодежи и спорта Украины
Донецкий Национальный Технический Университет
ВВЕДЕНИЕ
Существует большая группа живых существ, не имеющих клеточного строения. Эти существа носят названия вирусов (от лат. "вирус" — яд) и представляют неклеточные формы жизни. Вирусы нельзя отнести ни к животным, ни к растениям. Они исключительно малы, поэтому могут быть изучены только с помощью электронного микроскопа.
Вирусы способны жить и развиваться только в клетках других организмов. Вне клеток живых организмов вирусы жить не могут, и многие из них во внешней среде имеют форму кристаллов. Поселяясь внутри клеток животных и растений, вирусы вызывают много опасных заболеваний. Среди вирусных болезней растений известна мозаичная болезнь табака, гороха и других культур. У больных растений вирусы разрушают хлоропласты, и пораженные участки становятся бесцветными.
ИСТОРИЯ ОТКРЫТИЯ ВИРУСОВ
ПРИРОДА И ПРОИСХОЖДЕНИЕ ВИРУСОВ
Современные представления о вирусах складывались постепенно. После открытия вирусов Д. И. Ивановским (1892) их считали просто очень мелкими микроорганизмами, не способными расти на искусственных питательных средах. Вскоре после открытия вируса табачной мозаики была доказана вирусная природа ящура, а еще через несколько лет были открыты бактериофаги. Таким образом, были открыты три основные группы вирусов, поражающие растения, животных и бактерии. Однако в течение длительного времени эти самостоятельные разделы вирусологии развивались изолированно, а наиболее сложные вирусы — бактериофаги — долгое время считались не живой материей, а чем-то вроде ферментов. Тем не менее, уже к концу 20-х – началу 30-х годов стало ясно, что вирусы являются живой материей, и примерно тогда же за ними закрепились наименования фильтрующихся вирусов, или ультравирусов. Позже приставки отпали, и укоренилось ныне применяемое обозначение — вирусы, под которым объединили вирусы растений, животных и бактериофаги — бактериальные вирусы. В конце 30-х – начале 40-х годов изучение вирусов продвинулось настолько, что сомнения в живой их природе отпали, и было сформулировано положение о вирусах как организмах , Основанием для признания вирусов организмами явились полученные при их изучении факты, свидетельствовавшие, что вирусы, как и другие организмы способны размножаться, обладают наследственностью и изменчивостью, приспособляемостью к меняющимся условиям среды их обитания и, наконец, подверженностью биологической эволюции, обеспечиваемой естественным или искусственным отбором. Концепция о вирусах как организмах достигла своего расцвета к началу 60-х годов, когда было введено понятие “вирион” как вирусного индивидуума. Однако в эти же годы, ознаменовавшиеся первыми успехами молекулярной биологии вирусов, начался и закат концепции о вирусах как организмах, Уже тогда одновременно с введением понятия “вирион” были показаны, с одной стороны, отличия их строения от строения клеток и даже был введен термин “архитектура” вирионов. С другой стороны, были обобщены факты, указывавшие на совершенно отличный от клеток тип размножения, который некоторое время называли дизъюнктивной репродукцией, подчеркивая разобщенность синтеза генетического материала и белков вирусов. В докладе на 1-м Международном симпозиуме был сформулирован основной критерий отличия вирусов от других организмов: генетический материал вирусов является одним из двух типов нуклеиновых кислот, в то время как организмы имеют оба типа нуклеиновых кислот.
2.1 Размеры вирусов
Вирусы – это мельчайшие живые организмы, размеры которых варьируют в пределах примерно от 20 до 300 мм; в среднем они раз в пятьдесят меньше бактерий. Как уже говорилось, вирусы нельзя увидеть с помощью светового микроскопа (так как их размеры меньше полудлины световой волны), и они проходят через фильтры, которые задерживают бактериальные клетки. Следует также отметить, что вирусы не способны воспроизводить себя вне клетки-хозяина. Они находятся на самой границе между живым и неживым. И это лишний раз напоминает нам, что существует непрерывный спектр все возрастающей сложности, который начинается с простых молекул и кончается сложнейшими замкнутыми системами клеток.
Поведение
ВИРУСЫ КАК ВОЗБУДИТЕЛЬ ЗАБОЛЕВ АНИЙ У РАСТЕНИЙ
Распространение вирусов по растению
Вирусы, введенные в растение путем механической инокуляции, медленно распространяются по непроводящей ткани от первично зараженных клеток к соседним. Скорость распространения ВТМ составляет примерно 1 мм в день, а иногда и меньше. По-видимому, вирус, попавший в клетку, сначала в ней размножается, а затем уже проникает в соседние клетки по межклеточным канальцам, или плазмодесмам. Прежде чем ВТМ начинает двигаться из первично зараженной клетки в соседнюю, проходит несколько часов. Из одной клетки в другую могут мигрировать как интактные вирионы, так и вирусная РНК, с помощью электронной микроскопии вирионы были обнаружены в плазмадесмах. По тканям растения могут распространяться также и вирионы, неспособные к созреванию.
Когда вирус попадает в проводящую ткань либо из соседних паренхимных клеток, либо непосредственно вводится туда насекомым - переносчиком, он быстро движется сначала по жилкам, затем по черешку листа и, наконец, попадает в стебель. В принципе вирус может распространиться по всему растению, причем степень генерализации процесса зависит как от свойств вируса - хозяина. Большинство вирусов, переносимых механически, относится к числу гистологически “неограниченных” вирусов, а это означает, что они могут проникать почти во все ткани зараженного растения. Как правило, первыми атакуются вирусом активно растущие ткани и корни. Перенос вируса на значительные расстояния происходит главным образом по флоэме вместе с током пластических веществ, хотя вирусы могут мигрировать и по водопроводящей ткани растения - ксилеля. Вирусы могут перемещаться вместе с водным раствором органических веществ по членикам ситовидных трубок флоэмы, но это не обязательно приводит к заражению вирусом этих клеток. Однако некроз флоэмы при вирусных болезнях растений - отнюдь не редкое явление, доказательством чего может служить некроз этой ткани, вызываемый вирусом скручивания листьев картофеля. Перенос вирусов семенами наблюдается редко, а перенос пыльцой - еще реже. Особенности процесса морфогенеза цветка, очевидно, таковы, что они препятствуют проникновению вируса в гаметы. Вообще же клетки апикальных меристем, зараженных вирусом растений, как правило, содержат мало вируса, а иногда и вовсе свободны от вирусных частиц.
Степень распространения вируса по зараженному растению определяется ответной реакцией зараженных клеток на инфекцию, а сама эта реакция может быть весьма различной. К наиболее выраженным ее проявлениям относятся некротические поражения. В этом случае клетки погибают так быстро, что часто даже не успевают передать вирус соседним клеткам. Обнаружено, что при некротической вирусной инфекции в клетках повышается уровень полифенолоксидазы.
К наиболее выраженным проявлениям реакции растений на заражение вирусом относятся почти бессимптомные инфекции, обнаруживаемые лишь по очень слабым симптомам или благодаря тому, что вирус, не причиняющий явного вреда одним растениям, вызывает заболевания других. В этих случаях зараженные клетки повреждаются незначительно и, как правило, сохраняют способность к делению. Типичной реакцией клеток растений на заражение некоторыми вирусами является интенсивное их деление и даже опухолевая трансформация. Механизм опухолеродного действия вируса раневых опухолей пока остается неизвестным. Следует учитывать возможность совместного действия гормонов растений с вирусами (синергизм) в процессе индукции и стимуляции роста опухоли. Большая часть изменений обменных процессов наблюдаемых при вирусных болезнях растений, вероятно возникает в результате побочных воздействий, оказываемых вирусной инфекцией на процессы фотосинтеза, дыхания, регуляции роста, а также транспорта воды, пластических и других веществ. Нарушения регуляции роста приводят к морфогенетическим аномалиям самой различной значимости - от мозаичности листьев и цветков до некротических поражений и аномальной пролиферации побегов и образования опухолей. В местах некротических поражений часто накапливаются некоторые вещества типа скополетина - флуоресцирующего ароматического соединения, однако доказательства специфической роли вируса в биосинтезе этих веществ пока отсутствуют.
Вирусы всегда являются паразитами и поэтому вызывают у своих хозяев определенные симптомы того или иного вида заболевания. Вирусы вызывают и задержку роста растений, что впоследствии приводит к снижению урожая. Ряд серьезных заболеваний вызывают вирусы желтой мозаики турнепса (ВЖМТ), табачной мозаики (ВТМ), карликовой кустистости томатов и бронзовости томатов. Появление полосок на некоторых сортах тюльпанов также обусловлено вирусом, а ведь цветоводы продают эти тюльпаны, выдавая их за особый сорт. Вирусы растений, по-видимому, всегда относятся к РНК-содержащим вирусам.
Когда растение поражают вирусные заболевания, его рост заметно ухудшается, так что можно подумать, что растение содержится в неблагоприятных условиях и ему не хватает питательных веществ. Обычно вирусные заболевания растений путают с хлорозом. К тому же вирусные заболевания растений трудно распознать сразу.
Заболевания растений, вызываемые вирусами. Поражают представителей различных семейств цветковых растений, хвойных, папоротников, водорослей и грибов. Часто резко снижают урожай сельско - хозяйственных культур и его качество. Вирусные заболевания растений делят на мозаики и желтухи. Основной симптом мозаик — мозаичная (неравномерная) окраска листьев, обусловленная нарушениями в пластидном аппарате клеток ассимиляционной паренхимы листьев. Из болезней этой группы наиболее вредоносны: мозаика табака, мозаика и стрик томата, морщинистая и полосчатая мозаики картофеля, мозаика свёклы и др. Для желтух характерны: общий хлороз листьев; расстройство роста; скручивание, курчавость листьев; чрезмерное скопление в них углеводов, вызывающее их жёсткость и хрупкость. К желтухам относят желтуху свёклы, закукливание злаков, скручивание листьев картофеля и т.д. Из этой группы вирусных заболеваний растений исключены бывшие в ней ранее желтуха астр, столбур томата и др., вызываемые микоплазмоподобными возбудителями.
Мозаики легко передаются с соком больных растений во время пикировки рассады, при пасынковании, при соприкосновении больных и здоровых растений и лёгком взаимном травмировании их, например, при ветре, иногда через семена, а также сосущими насекомыми. Перенос вируса при этом происходит чисто механически. Желтухи распространяются преимущественно насекомыми-переносчиками, главным образом цикадами. Передача вирусов происходит биологически после предварительного размножения вируса в теле насекомого. Переносчиками вирусных заболеваний растений могут быть и растительноядные клещи, нематоды, низшие грибы. Возможна передача вирусов повиликой. Почти все вирусные заболевания растений легко передаются потомству при вегетативном размножении, прививках.
Вирусы зимуют в растениях, в их отмерших остатках, в переносчиках, в посевном и посадочном материале. На скорость размножения вирусов в растительных тканях и на проявление симптомов болезни большое влияние оказывают возраст растений (наиболее восприимчивы молодые растения), условия их питания и другие факторы внешней среды.
Механизмы передачи вирусов растений
Механическая передача вирусов растений на поверхность листа может быть осуществлена в эксперименте со многими вирусами, однако маловероятно, чтобы такой путь передачи был основным способом распространения вирусов растений в естественных условиях. Одним из немногих исключений в этом отношении является ВТМ. Передачу почти любого вируса можно осуществить прививкой. Хотя этот способ заражения используется главным образом при экспериментальных исследованиях, он может играть значительную роль в распространении вирусных болезней плодовых деревьев и декоративных кустарников. Паразитное растение повилика, гаустории которой внедряются в стебли растений - хозяев (таким образом устанавливается живая связь между сосудистыми системами растения - хозяина и паразита), служит полезным инструментом для изучения передачи вирусов новым хозяевам. Однако очевидно, что такой механизм передачи вирусов не может быть основным способом их распространения в природе.
22 августа 2013
- 4202
- 3,5
- 0
- 0

Спонсор конкурса — дальновидная компания Thermo Fisher Scientific. Спонсор приза зрительских симпатий — фирма Helicon.
Чем болеют растения?
Для начала несколько слов о том, от чего, собственно, специалистам приходится защищать сельскохозяйственные растения. Причинами заболевания растений могут быть как факторы среды (летняя засуха или зимние морозы, недостаток питательных веществ в почве или их избыток и т.п.), так и различные паразитические организмы (бактерии, вирусы, грибы, круглые черви (нематоды) и даже другие растения).
Грибы, бесспорно, являются основными патогенами культурных растений. Известно, например, что из 162 серьёзных заболеваний в Центральной Европе 135 (83%) вызываются грибами [2]. Фитопатогенные грибы — многочисленная группа; их описано свыше 10 000 видов, различных по систематическому положению, степени паразитизма, специализации и т.д. [3]. Они широко распространены в природе и при благоприятных для их развития условиях наносят значительный урон урожаю и сельскохозяйственным продуктам при хранении. Даже самые осторожные оценки говорят об уничтожении болезнями 10–20% потенциального урожая; без контрмер масштабы этих потерь резко возросли бы [2].
Именно о проблемах диагностики болезней растений, вызываемых фитопатогенными грибами, пойдёт речь в данной статье.
Врага надо знать в лицо
Зачем же нужно, с одной стороны — обнаружение, а с другой — быстрое и точное (желательно — до вида, или даже расы) определение фитопатогенных грибов?
На данный момент самым распространённым методом борьбы с фитопатогенными грибами является обработка растений фунгицидами. Понятно, что невозможно защитить культуры от всех возможных потенциальных угроз: это и сложно, и экономически невыгодно, да и для окружающей среды далеко не полезно. Именно поэтому важно знать, желательно — своевременно, с чем именно придётся бороться. Чем раньше обнаружена болезнь, тем больше шансов, что, приняв соответствующие меры, удастся её победить. Это верно для заболеваний как человека, так и растений. Кстати, точное определение вида грибов важно ещё и в довольно неожиданной области — реставрации деревянных строений — поскольку используемые там антисептические меры также очень сильно зависят от типа поражения [4].
Кроме этого, идентификация фитопатогенных грибов необходима для изучения их таксономии и эволюции, их взаимоотношений с растениями-хозяевами, генетических основ восприимчивости и устойчивости растений, что, в конечном счете, должно помочь в разработке способов борьбы с патогенами и в селекции растений, невосприимчивых к болезням [5].
И, наконец, крайне важна сертификация зерна и посадочного материала в рамках карантинных программ. Известно, что фитопатогенные грибы могут распространяться многими путями — как естественными (с током воздуха, водой, насекомыми, животными), так и при помощи человека, перевозящего заражённые растения или их части не только между различными странами, но и между континентами. Зачастую такое перемещение приводит к неожиданному и масштабному распространению заболеваний.
Например, пузырчатая ржавчина (Cronartium ribicola) была эндемична для Альп и востока России. Этот паразит, в цикле развития предполагающий обязательную смену хозяев, обитает круглый год на пятихвойных соснах, а летом поражает листья смородины; ни в одном из исходных ареалов он не причинял серьёзного ущерба. Однако веймутова сосна, завезённая в начале XVIII века из Америки в ряд областей Европы, оказалась крайне восприимчивым хозяином для данного гриба. За счёт этого распространившаяся инфекция причинила большой вред культурам смородины и высаженным веймутовым соснам, а в 1909 году была завезена с их рассадой в Америку, где встретила многочисленных хозяев для обеих фаз развития. Здесь стали страдать, прежде всего, лесообразующие пятихвойные сосны. Поэтому, чтобы разорвать инфекционную цепь паразита с обязательной сменой хозяев, пытаются уничтожать дикорастущие виды смородины [2].
Ещё один показательный пример: возбудитель голландской болезни вяза (Ophiostoma ulmi) уже в XX столетии был занесён из континентальной Европы в Северную Америку. Начиная примерно с 1970 г., после того, как он был завезён в Великобританию, он успел уничтожить половину английских вязовых насаждений [2]. Теперь этот вид встречается и в России.
Для того чтобы избежать подобного впредь, созданы списки карантинных организмов, и при перемещении растений или их семян между странами (или даже частями одной страны) обязательно проводится их обследование.
Как только что было показано, идентификация фитопатогенных грибов крайне важна, возник вопрос — каким образом она производится?
Наиболее простой способ — это идентификация патогена по внешним признакам заболевания (симптомам), то есть по тому воздействию, которое он оказывает на поражённое растение [6]. Но здесь проблема в том, что к одним и тем же повреждениям растения-хозяина могут приводить совершенно разные микроорганизмы, отличающиеся разной устойчивостью к фунгицидам, вредоносностью и другими характеристиками. Как пример, здесь можно привести три листовые пятнистости пшеницы (рис. 1).

Рисунок 1. Листовые пятнистости пшеницы. Слева — септориоз листьев пшеницы (возбудитель — Mycosphaerella graminicola). По центру — септориоз листьев и колоса пшеницы, проявление на листьях (возбудитель — Phaeosphaeria nodorum). Справа — жёлтая пятнистость пшеницы (возбудитель — Pyrenophora triticirepentis). Обратите внимание: несмотря на то, что это разные заболевания, поражения листьев очень похожи.
Ещё одна проблема заключается в том, что далеко не все заболевания проявляются сразу же после заражения растения. Например, возбудитель пыльной головни ячменя (Ustilago nuda) обычно проникает во время цветения пшеницы в формирующуюся зерновку. Гриб не препятствует формированию зародыша, само зерно развивается нормально, ничем внешне не отличаясь от здорового. Мицелий зимует в зерновке. Весной одновременно с прорастанием семян происходит и рост мицелия, который по мере роста растения распространяется по различным его органам. Проявляется заболевание только в период колошения. При этом разрушаются все части колоса, превращаясь в чёрную споровую массу, после распыления которой остаются лишь ости и стержень колоса (рис. 2) [8].

Рисунок 2. Пыльная головня ячменя: поражённое соцветие со спорами
Стандартный для фитопатологов подход при определении фитопатогенных грибов — это выделение их в чистую культуру на какой-либо питательной среде, получение характерных образований (чаще всего это, конечно, спороношения) и затем идентификация гриба под микроскопом.
Но здесь возникают определённые трудности. Основная из них заключается в том, что далеко не все паразитические грибы возможно культивировать на искусственных питательных средах: многим требуется наличие живых тканей растения-хозяина, либо присутствие других представителей сложного сообщества [10]. Но даже если гриб удаётся выделить в культуру, следующий вопрос — это то, сколько времени понадобится, чтобы добиться от него появления спороношения. Например, возбудитель белосоломенной болезни пшеницы и ржи (Gibellina cerealis), хотя и хорошо культивируется, даёт спороношение только после четырёх–пяти недель роста. Естественно, что меры по борьбе с патогеном необходимо принимать сразу после его обнаружения, а не через месяц, когда может оказаться, что спасать уже нечего.
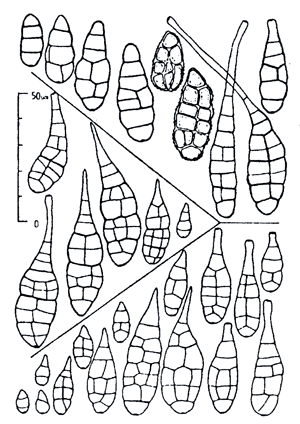
Рисунок 3. Сравнение конидий типовых образцов Alternaria longipes (вверху), Alternaria tenuissima (в центре), Alternaria alternata (внизу). Видно, что на основе сравнения только формы конидий этих трёх видов однозначно различить их крайне сложно. При идентификации видов в данном случае специалист использует не только форму конидий, но и другие признаки (например, способ образования конидий, их взаимное расположение и т.п.).
И даже с определением тех фитопатогенных грибов, спороношения которых получить сравнительно просто, могут возникать сложности. К примеру, идентификация многих микромицетов сопряжена с рядом трудностей, таких как сходство морфологических характеристик разных видов и одновременно внутривидовая вариабельность признаков. Несмотря на внешнее сходство, возбудители могут значительно отличаться по патогенности, токсигенности, степени специализации, генетике взаимоотношений с растением-хозяином, вредоносности, чувствительности к фунгицидам и т.д. То есть разные виды обладают совершенно разными экологическими особенностями и хозяйственной значимостью [12]. Хорошим примером здесь является определение различных видов рода Alternaria (рис. 3). Очевидно, что для идентификации до вида нужны достаточно широкие познания в данной области и немалый опыт работы с исследуемым фитопатогеном.
Ещё один способ, пригодный для обнаружения некоторых фитопатогенных грибов, заключается в смыве с субстрата, фильтрации и микроскопическом определении (и даже подсчёте, что даёт количественные данные) их спор. Чаще всего, таким способом оценивается количество грибных спор в зерне или в почве. Несмотря на то, что идентификация до вида на основании одних только спор чаще всего затруднена, этот способ широко применяется, а для анализа получаемых при помощи микроскопа изображений разрабатываются специальные компьютерные программы [14]. Например, таким образом определяют заражённость зерна возбудителем твёрдой головни (Tilletia caries) (рис. 4) [15]. Несмотря на использование компьютерных технологий, этот метод весьма трудоёмок и не подходит для исследования большого количества образцов.

Рисунок 4. Зерновки, поражённые твёрдой головнёй пшеницы
Молекулярная биология на службе фитопатолога
Во всех описанных случаях на помощь исследователям могут прийти широко развивающиеся в последнее время молекулярные методы анализа. Сейчас в основе большинства из них лежит применение ELISA (enzyme-linked immunosorbent assay, иммуноферментный анализ) [11], либо ПЦР (полимеразная цепная реакция, polymerase chain reaction) [17].
Иммуноферментный анализ состоит из двух основных этапов: иммунной и ферментативной реакций. Иммунная реакция заключается в специфическом связывании характерного для данного микроорганизма антигена с диагностическим антителом. Ферментативная реакция необходима для обнаружения этого связывания. Как правило, она сопровождается изменением цвета, причём степень этого изменения может быть использована для определения количества присутствующего антигена.
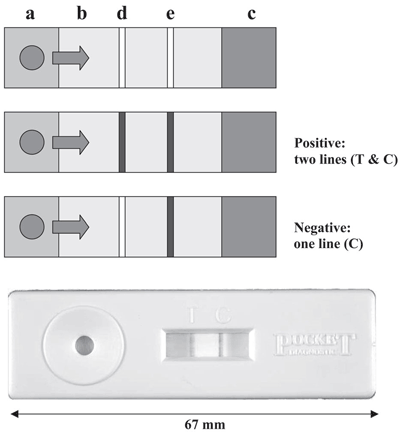
Рисунок 5. Прибор CSL Pocket Diagnostic TM lateral flow immunodiagnostic kit. Растительный экстракт помещается на площадку (a), которая содержит латексные шарики, покрытые специфическими антителами; смесь мигрирует вдоль мембраны (b) к абсорбирующей поверхности (c). При этом имеющиеся в растворе целевые антигены связываются со специфичными антителами на латексных шариках. Мембрана содержит полосу антител, отличающихся необходимой специфичностью (измерительную полосу) (d) и полосу других антител, которые связываются с первыми антителами (контрольную полосу) (e). Латексные шарики, содержащие связанный антиген, задерживаются в тестовой зоне, давая видимую линию, тогда как излишние латексные шарики, которые не содержат антигена, задерживаются в контрольной зоне, показывая, что анализ работает. Наличие двух линий соответствует положительному результату (positive), наличие только одной линии (контрольной) говорит о негативном результате (negative).
Основанные на иммуноферментном анализе методы широко применяются для обнаружения вирусов (в том числе поражающих растения) и значительно реже — для идентификации грибов и бактерий. Основной причиной этого является трудность получения антител с необходимой специфичностью: строение клеточных стенок грибов и бактерий гораздо сложнее, чем вирусного капсида, к тому же может изменяться в ходе их жизненного цикла. В результате получаемые антитела могут оказаться специфичны как сразу к большой группе видов, так и исключительно к отдельным жизненным формам данных микроорганизмов. Тем не менее, основанные на ELISA методы идентификации фитопатогенных грибов всё же разрабатываются: например, существует метод идентификации спор уже упоминавшейся в данной статье твёрдой головни [19].
ПЦР — это ферментативная реакция, в результате которой происходит накопление большого количества копий какого-либо не слишком большого (чаще всего, 200–1500 пар нуклеотидов) фрагмента ДНК. Так как ДНК любого организма содержит как вариабельные (отличающиеся даже у близкородственных организмов), так и консервативные (сходные у эволюционно далёких видов) участки, возможно на основе выбора диагностического участка варьировать специфичность протекающей реакции.
Таким образом, данный метод позволяет обнаруживать последовательности нуклеиновой кислоты, специфичные для конкретного организма или группы сходных организмов и, тем самым, выявлять его (их) присутствие в анализируемой пробе. Методы, основанные на ПЦР, позволяют идентифицировать патогенные виды как в чистой культуре, так и непосредственно в растительном материале, минуя этап изоляции грибов [20]. Как пример, здесь приведены результаты ПЦР, разработанной для идентификации грибов рода Pyrenophora (рис. 6), представители которого являются возбудителями жёлтой пятнистости злаков, в частности — пшеницы (рис. 1).
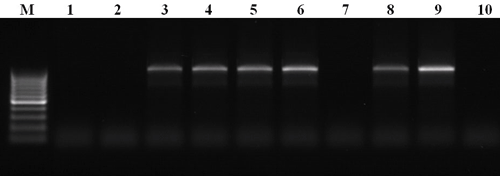
Рисунок 6. Разделённые при помощи электрофореза продукты ПЦР, разработанной для идентификации грибов рода Pyrenophora. М — маркер, представляющий собой набор фрагментов ДНК известного размера, 1–10 — ДНК, выделенная из различных образцов листьев пшеницы, поражённых листовыми пятнистостями. Здесь продукт реакции (фрагмент ДНК известного размера) должен наблюдаться только в том случае, если в образце присутствует ДНК целевого организма, а именно — гриба рода Pyrenophora. В итоге видно, что растения под номерами 3–6, 8 и 9 больны жёлтой пятнистостью, а остальные — каким-либо другим внешне схожим заболеванием.
Существует достаточно много модификаций метода ПЦР, большинство из которых применяется в изучении возбудителей болезней растений. Например, RAPD и RFLP анализы используются для уточнения родственных связей между различными грибами; ПЦР, специфичная для ДНК представителей отдельных родов или видов — для идентификации фитопатогенов (в том числе — в форматах nested и multiplex); ПЦР с регистрацией в режиме реального времени (real-time PCR) — для определения количества присутствующей целевой ДНК.
Рассмотрим подробнее один из самых перспективных методов на основе ПЦР — ПЦР с регистрацией в режиме реального времени (рис. 7). В отличие от большинства других форматов ПЦР, он позволяет не только констатировать факт присутствия ДНК целевого патогена, но и измерить её количество. В качестве примера здесь приведено определение в двух образцах количества ДНК ещё одного возбудителя листовой пятнистости.
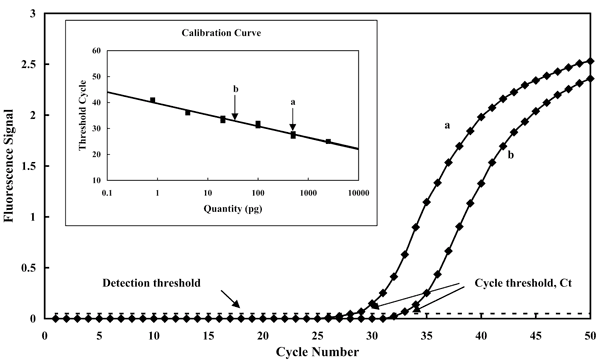
Интересно применение данного метода для анализа заражённости зерна твёрдой головнёй (рис. 4): при наличии соответствующих калибровочных графиков возможно получение результатов в виде числа спор, имеющихся в образце [7].
Ложка дёгтя в бочке мёда
Хотя преимущества и перспективы применения молекулярных методов идентификации сложно переоценить, на пути их практического использования имеется целый ряд трудностей. Несмотря на универсальность методов при конечном анализе, для их разработки и проверки требуется достаточно много времени и немалая экспериментальная база. Основной проблемой здесь является отсутствие возможности чисто теоретически оценить специфичность разрабатываемых методов.
Ну и самая большая проблема всех описанных в данной статье методов — это цена, ограничивающая их широкое применение в условиях небогатых российских хозяйств.
Несколько слов о будущем
Несмотря на все имеющиеся проблемы, молекулярные методы анализа интенсивно развиваются (о чём можно судить хотя бы по числу публикаций на соответствующие темы, которое с каждым годом становится всё больше). Старые методы постоянно совершенствуются, в то же время разрабатываются новые (например, метод биочипов [21] и секвенирование следующего поколения [22]), а цена одного анализа становится всё ниже. Поэтому можно надеяться, что не за горами то время, когда все упоминавшиеся в данной статье методики и их более совершенные аналоги действительно найдут широкое применение и облегчат жизнь фитопатологов и агрономов.
Читайте также: