Эндосимбиоз прокариот и эукариот
Добавил пользователь Владимир З. Обновлено: 29.01.2026
Дарвин задокументировал естественный отбор среди зябликов на Галапагосских островах. Обладая новыми знаниями, Дарвин предложил свою теорию эволюции. Он утверждал, что эволюция был по существу спуск с модификация. Со временем ученые адаптировали определение эволюции. Эволюция теперь рассматривается как естественный отбор, ведущий к потомству с модификациями. Нет сомнения, что теория Дарвина заложила основу для изучения эволюции. Однако многие не знают об эндосимбиозе. Эта теория касается одноклеточных организмов, которые бродили по Земле задолго до появления сложных многоклеточных вьюрков. Ведь эндосимбиоз имеет первостепенное значение в понимании клеточная биология и спуск с модификацией.
Что такое теория эндосимбиоза?
«Эндо» относится к чему-то внутреннему. «Симбиоз» определяется как взаимодействие двух организмов. Существуют различные типы симбиотических отношений; мутуализм, комменсализм и паразитизм. Мутуализм - это симбиотические отношения, приносящие пользу обоим видам. Комменсализм приносит пользу только одному виду, в то время как другой остается неизменным. Кроме того, паразитизм приносит пользу только одному виду. Но в этих отношениях один вид страдает от другого. Следовательно, эндосимбиоз - это когда симбиотический организм живет внутри другого организма.
Эндосимбиоз относится к видам, но теория эндосимбиоза к видам не относится. Вместо этого теория касается органелл. Линн Маргулис, гипотетический эндосимбиоз в отношении митохондрий, хлоропластов (в клетках растений) и базальных тел (часть жгутиков). Ее адаптация к этой теории, впервые предложенная Иваном Валленом в 1920-х годах, заявила, что эти органеллы когда-то были одноклеточными организмами, и они были поглощены ранними эукариотическими клетками.
Прокариотические и эукариотические клетки
Прокариотические клетки и эукариотические клетки имеют мало общего и во многом отличаются друг от друга. Оба типа клеток имеют схожие особенности структуры, цитоскелета и цитоплазмы, заполняющей внутриклеточное пространство. Прокариоты и эукариоты также имеют рибосомы - органеллы, ответственные за синтез белков. Помимо этих немногих сходств, эти типы клеток сильно различаются по размеру, структуре и составу.
Прокариотические клетки
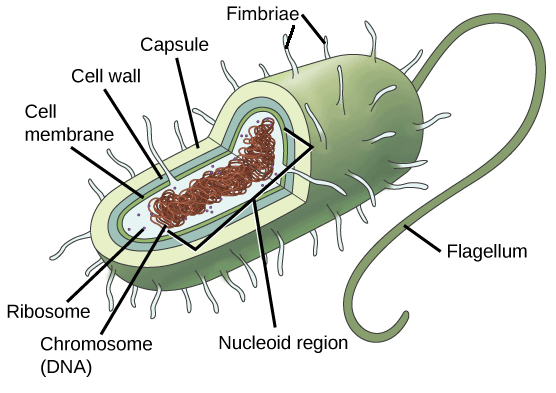
Типичная прокариотическая клетка. Эти клетки не состоят из многих компонентов. Источник изображения: Khan Academy
Эукариотические клетки
Эукариотические клетки намного больше прокариотических клеток, от 10 до 100 микрометров в диаметре. Следовательно, в них больше места для органелл. У эукариот есть эндоплазматическая сеть, Гольджи, микротрубочки, лизосомы, митохондрии и хлоропласты (у автотрофов) и многие другие. Эукариотические клетки составляют многоклеточный организм, такой как животные и растения, и размножаются посредством митоза (бесполое размножение). Мейоз происходит в результате полового размножения и допускает генетические вариации между разными поколениями. Более того, можно наблюдать некоторые черты прокариот у эукариот, за исключением более крупных масштабов. Например, у них больше ДНК. Эукариотическая ДНК обвивает гистоновые белки, обеспечивая правильную экспрессию генов. Экспрессия генов варьируется, потому что часть ДНК представляет собой 10-нанометровое волокно с большей экспозицией, тогда как остальная часть - 30-нанометровое волокно с меньшей экспозицией. У эукариот также есть более крупные рибосомы для более разнообразной экспрессии генов и более крупные жгутики, которые продвигают более крупные клетки.
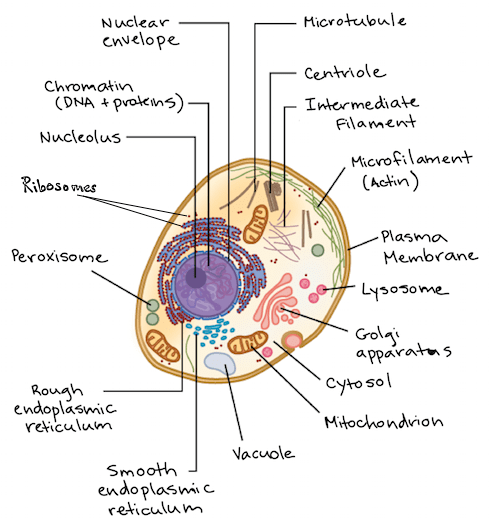
Типичная эукариотическая клетка. У эукариот больше органелл по сравнению с прокариотами. Источник изображения: Khan Academy
Доказательства, подтверждающие теорию эндосимбиотиков
Согласно теории Маргулиса, митохондрии и хлоропласты - это одноклеточные организмы. Следовательно, все они были меньше ранних эукариотических клеток, были более простыми и содержали меньше ДНК. На основе этих органелл следует доказательство теории эндосимбиотиков и их роль в эволюции.
Митохондрии
Митохондрии вырабатывают для клетки аденозинтрифосфат (АТФ). АТФ содержит большую потенциальную энергию в своих фосфодиэфирных связях, высвобождаемых при образовании аденозиндифосфата (АДФ). Белки служат для поддержки и катализирования метаболических процессов. Одни только митохондрии могут кодировать до 63 белков, но содержат порядка 2000 белков (Zimorski, V. et al., 2014). Есть несоответствие количества белков. Как органелла может кодировать только 63 белка, но потенциально может создать 2000? Исследователи Zimorski et al. обязаны этим несоответствием переносу эндосимбиотического гена.
Эндосимбиотический перенос гена
Репозиционирование генов происходит во время эндосимбиотического переноса (Zimorski, V. et al., 2014). Во время этого процесса гены перемещаются из митохондрий в хромосомы клетки-хозяина. Цитозольные белки экспрессируют многие гены, связанные с митохондриями. В результате эти цитозольные белки дублируют сигнальные молекулы для митохондрий (Zimorski, V. et al., 2014). Поскольку экспрессия митохондриальных генов происходит вне митохондрий, ядерный геном расширяется (Zimorski, V. et al., 2014).
Биоэнергетика в эволюции эукариот
Различные митохондриальные гены имеют разное происхождение. Например, некоторые гены содержат высокие концентрации аденина и тимина. Поэтому ученые прослеживают эти гены до протеобактерий, богатых аденином и тимином (Zimorski, V. et al., 2014). Другие ученые проследили происхождение эукариот до более простых архей. В любом случае, кристы митохондрий, производящие энергию, обеспечивают работу эукариотических клеток.
Хотя ученые все еще собирают научные факты о происхождении митохондрий, есть несколько широко обсуждаемых гипотез о дополнительных ролях, которые митохондрии играют в клетке. Обзор исследований, проведенный Линчем и Мариновым, собрал данные о взаимосвязи между митохондриями и характеристиками клеток. По мере увеличения площади поверхности клетки в микрометрах площадь поверхности митохондрии также увеличивается (Lynch, M., & Marinov, GK, 2017). Кроме того, увеличенная площадь поверхности митохондрии коррелирует с увеличением количества комплексов АТФ-синтазы, расположенных на органелле (Lynch, M., & Marinov, GK 2017). АТФ-синтаза - это белок, ответственный за создание протонного градиента, который способствует фосфорилированию АТФ до АДФ.
Взаимоотношения
Поскольку в клетках с большим объемом (эукариотах) больше митохондрий и больше молекул АТФ, больше АТФ доступно для метаболических процессов. Следовательно, эукариотические клетки становятся более сложными с большим количеством биоэнергетических мембран. Рибосомы во многом связаны со сложностью эукариотических клеток. Они собирают белки, необходимые для экспрессии генов. Но с молекулами АТФ работа отдельных генов является обычным делом, а эволюционные модификации генов происходят чаще (Lynch, M., & Marinov, GK 2017). Рибосомы и митохондрии являются индикаторами способностей клетки.
Митохондрии имели мутуалистические отношения с клеткой-хозяином, прежде чем стать зависимыми от клетки-хозяина. Мартин и Мюллер (1998) выдвинули гипотезу о преимуществах этих отношений. Они считали, что митохондрии снабжают клетку побочными продуктами водорода, необходимыми для метаногенеза (Martin W, Müller M., 1998). В свою очередь, клетка-хозяин обеспечивала митохондрии необходимыми ресурсами (Martin W, Müller M., 1998). В противном случае митохондрии получили бы эти ресурсы из окружающей среды.
Хлоропласты
Хлоропласты имеют общие характеристики с митохондриями. Как и митохондрии, они имеют кольцевую ДНК и подвергаются бинарному делению. Кроме того, гены хлоропластов интегрируются в ядерный геном ранних эукариотических клеток посредством переноса эндосимбиотических генов. Эти факторы совпадают с характеристиками прокариот. Но у хлоропластов есть дополнительные особенности, которые еще больше подтверждают эндосимбиотическую теорию.
Филогенетический анализ эндосимбиоза хлоропластов
Чтобы обнаружить доказательства эндосимбиоза в хлоропластах, ученые наблюдали гены, кодирующие хлоропласты. Связи между семействами ферментов и генами-гомологами позволяют проводить филогенетический анализ. Ученые исследовали ферменты, участвующие в производстве жирных кислот и липидов, входящих в состав мембран хлоропластов. Результаты приведены ниже.
Тип 1 Дерево
Ферменты хлоропластов произошли от цианобактерий (Sato, N., 2020). Белки, такие как галактозилтрансфераза, участвуют в перемещении углеводов. Белки хлоропластов выделены из монолитной группы (зеленые бактерии и цианобактерии). Но хлоропласты - это органеллы, а цианобактерии - прокариоты. Это дерево лучше всего иллюстрирует расхождение эндосимбиоза и ферментов хлоропластов (Sato, N., 2020). Вероятно, их различия в первую очередь связаны с событием эндосимбиоза (Sato, N., 2020).
Тип 2 Дерево
В дереве типа 2 и белки хлоропластов, и цианобактерии представляют собой монолитную группу. Эта группа отошла от зеленых бактерий. Между белками хлоропластов и цианобактериями также есть синапоморфия. Сато заявляет, что эта синапоморфия является «сестринской группой» фермента. Сестринские группы похожи, но не идентичны.
Тип 3 Дерево
Дерево типа 3 состоит из двух отдельных монолитных групп. В этом случае у каждой группы своя синапоморфия. Другими словами, существует синапоморфия между хлоропластами и белками хлоропластов зеленых бактерий. Синапоморфия между цианобактериями и другими бактериями означает отсутствие ферментов хлоропластов.
Это дерево контрастирует с деревьями типа 1 и типа 2, поскольку хлоропласты и цианобактерии расположены дальше друг от друга. Сато предполагает, что белки хлоропластов произошли от гомологов, а не от цианобактерий (2020). Следовательно, он подразумевает, что у них есть общий предок, гены которого остаются неизменными с течением времени.
Тип 4 Дерево
В этом филогенном дереве хлоропластные белки ответвляются от эукариотической монолитной группы. Между тем прокариотическая группа отличается. Если эукариоты и белки хлоропластов очень тесно связаны, то белки хлоропластов, вероятно, произошли от эукариот (Sato, N., 2020).
Итоги
Существует множество свидетельств происхождения хлоропластов в зависимости от того, какие гены наблюдают ученые. Однако ученые обнаружили тип 1 при анализе рибосомной РНК (рРНК), домашних белков в красных водорослях и консервативных белков в красных водорослях и цианобактериях (Sato, N., 2020). Все эти факторы подтверждают тот факт, что хлоропласт мог выжить как свободноживущий организм, что дополнительно подтверждает эндосимбиотическую теорию.
Четыре модели филогении указывают на сложность хлоропластов и возможный эндосимбиоз. Некоторые ученые считают, что хлоропласты претерпели множественные процессы эндосимбиоза, чтобы полностью интегрироваться в эукариот (Jensen, PE, & Leister, D., 2014). Независимо от того, как происходит эндосимбиоз, хлоропласт служит цели обеспечения клетки необходимыми химическими соединениями, которые создают химическую энергию (Jensen, PE, & Leister, D., 2014). Технические трудности ограничили изучение многих белков, участвующих в фотосинтезе (Jensen, PE, & Leister, D., 2014). Несмотря на эти трудности, ученые обнаружили новые ретроградные сигналы.
Новые ретроградные сигналы
Ретроградные сигналы связаны с сигналами от хлоропластов или митохондрий (Jensen, PE, & Leister, D., 2014). Эти сигналы взаимодействуют с ядром, чтобы модулировать экспрессию генов (Jensen, PE, & Leister, D., 2014). Многие из этих сигналов также не были обнаружены из-за технических трудностей. Но они действительно подразумевают другую адаптацию этих органелл после эндосимбиоза (Jensen, PE, & Leister, D., 2014). Уменьшение геномов органелл происходит после эндосимбиоза. В результате органеллы должны взаимодействовать с другими клеточными компонентами, и эти органеллы исходят из мутуалистических отношений к зависимым отношениям с остальной частью клетки. Эти сигналы часто являются результатом побочных продуктов метаболизма (Jensen, PE, & Leister, D., 2014). Они также показывают, что эволюция хлоропластов и даже митохондрий зависит от других клеточных компонентов.
Эндосимбиотическая теория и эволюция
Эндосимбиоз касается органелл. И митохондрии, и хлоропласты несут метаболические процессы (дыхание и фотосинтез). Ученые часто изучают экспрессию генов в надежде узнать больше о том, как функционируют клетки. Однако эндосимбиоз подчеркивает важность метаболизма и его связь с экспрессией генов.
Микроэволюция
Микроэволюция относится к эволюции, происходящей ниже уровня вида (O'Malley, MA, 2015). Эволюция является результатом мутаций в геноме вида. На клеточном уровне могут происходить молчащие мутации, но они могут влиять на сложные белковые структуры, необходимые либо для фотосинтеза, либо для дыхания. О'Мэлли ссылается на случаи, когда эндосимбиоз не имеет прочных оснований в микробиологии. Но его предполагаемая важность в эукариогенезе дополнительно подтверждает одну из ключевых идей эволюции: происхождение с модификацией. Следовательно, ученые могут применять идеи эндосимбиоза для изучения заболеваний, связанных с этими органеллами.
Заболевания митохондрий
Некоторые заболевания, поражающие людей, приводят к митохондриальная дисфункция. Диабет, рак, мышечная дистрофия и даже болезнь Альцгеймера являются вторичными заболеваниями митохондриальной дисфункции. Примечательно, что каждый 5,000 человек имеет генетическую дисфункцию, которая влияет на митохондрии.
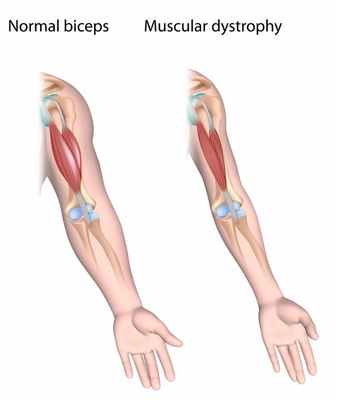
На изображении выше сравниваются бицепсы человека с мышечной дистрофией и человека без заболевания. Митохондрии влияют на размер мышц. Следовательно, без достаточного объема митохондрий мышцы атрофируются. Источник изображения: medlineplus.gov
Митохондрии когда-то были свободноживущими организмами и до сих пор имеют общие характеристики со своими бактериями и родственниками архей. Например, ученые иногда помещают прокариот в полноценную среду во время метаболической дисфункции. Они делают это, чтобы обеспечить наилучшие условия для метаболизма. Понимание оптимальных условий для митохондриальной активности может быть лекарством от митохондриальной дисфункции.
Заболевания, поражающие хлоропласты
Заболеваний, поражающих хлоропласт непосредственно у растений, не так много, но вирусы делать. Однако ученые обнаружили, что хлоропласт играет ключевую роль в синтезе вирусная РНК. Известно, что белки хлоропластов взаимодействуют с некоторыми вирусными белками. Фактически, они даже привлекают вирусные компоненты в хлоропласт, где происходит репликация РНК.
Хлоропласты, обеспечивающие сайт репликации вирусной РНК, позволяют создавать бактериофаги. Фаги представляют угрозу для бактерий, а некоторые бактерии полезны. Как и в случае с митохондриями, понимание метаболизма хлоропластов может быть полезным. Если ученые найдут оптимальные условия для репликации вирусной РНК, они смогут продолжить изучение методов предотвращения репликации. Даже в этом случае профилактика должна затрагивать только репликацию вирусной РНК, не влияя на метаболизм бактерий.
Возможности теории эндосимбиотиков
Ученые сталкиваются с препятствиями при изучении отдельных органелл из-за технологических ограничений. Следовательно, эндосимбиоз до сих пор остается теорией. Исследования может или не может в дальнейшем подтвердить эндосимбиоз с расширением научных знаний. Но, обладая знаниями, собранными к настоящему времени, ученые могут изучать метаболический аспект нарушений и работать над их борьбой с помощью лечения, помимо тех, которые направлены на изменение генома.
Эндосимбиоз не только обладает большим потенциалом, но и показывает, как происходит научное знание. Наука не есть истина. Скорее, наука — это процесс получения истины и сбора знаний, окружающих ее. Ученые не обладают достаточными знаниями о функциях хлоропластов и митохондрий до и после эндосимбиоза. В результате влияние, которое эти органеллы оказывают на эукариотическую клетку, по большей части только предполагается. На данный момент использование метаболических знаний для лечить болезни все еще является новым и непроверенным. Но эта гипотеза имеет достаточно оснований, чтобы доказать ее несостоятельность.
Рекомендации
Мартин В., Мюллер М., 1998. Гипотеза водорода для первого эукариота. Природа. 1998;392:37–41. DOI: 10.1038/32096.
Эндосимбиоз прокариот и эукариот
Эндосимбиоз прокариот и эукариот
• Митохондрии и хлоропласты образовались при интеграции прокариот в цитоплазму эукариотических клеток где они стали постоянными симбиотическими резидентами
• У некоторых видов растительных культур ризобии образуют клубеньки, и атмосферный азот может превращаться в биологически усваиваемый аммиак
• Развитие и жизнеспособность гороховой тли зависят от эндосимбиоза с бактериями Buchnera
Под термином эндосимбиоз понимают такие взаимоотношения между двумя симбиотическими организмами, при которых один организм существует внутри другого. Обычно такие взаимоотношения выгодны для одного или для обоих организмов. Эндосимбиотическое сосуществование с прокариотами определило многие пути эволюции эукариот. В первую очередь, следует отметить эндосимбиоз, в результате которого в эукариотических клетках появились митохондрии и хлоропласты. Они представляют собой органеллы, в которых происходят процессы окислительного фосфорилирования, в результате чего образуется АТФ. Без митохондрий эукариотические клетки зависели бы от анаэробного гликолиза, который, по сравнению с аэробным дыханием, является крайне малоэффективным. За счет дыхания образуется в 15 раз больше молекул АТФ, чем при гликолизе. Неудивительно поэтому, что клеткам таких органов, как сердце и скелетные мышцы, которые потребляют много энергии, необходимо много митохондрий. В клетках растений хлоропласты превращают энергию солнечного света в АТФ в ходе процесса, называемого фотосинтезом.
Линн Маргулис привела убедительные доводы в пользу того, что митохондрии и хлоропласты представляют собой потомков микробов, которые вначале вели самостоятельное существование, но в какой-то момент начали симбиоз с клетками эукариот. Существует много доказательств в пользу такого эндосимбиоза. Эти органеллы образуются только из предсуществующих митохондрий и хлоропластов, поскольку ядерные гены кодируют лишь часть белков, входящих в их состав, — остальные кодируются генетическим материалом самой органеллы. Митохондрии и хлоропласты имеют собственный геном, напоминающий геном прокариот. Он содержит кольцевые молекулы ДНК, не связанные с гистонами, которые способны делиться и реплицироваться независимо от репликации клетки хозяина.
Наиболее убедительное доказательство прокариотического происхождения митохондрий заключается в том, что гены рибосомальной РНК обнаруживают явное прокариотическое, а не эукариотическое происхождение. Секвенирование митохондриального генома простейшего Reclinomonas americana показало, что он содержит около 70 генов, почти половина из которых кодирует компоненты аппарата трансляции. Остальные кодируют ферменты, принимающие участие в энергетических процессах митохондрий.
Филогенетический анализ рибосомальных белков, цитохромоксидазы и NADH дегидрогеназ митохондрий R. Americana продемонстрировал близкое родство между митохондриями и представителями группы а-протеобактерий. Большинство бактерий этой группы существуют совместно с клетками эукариот или как симбионты, или как паразиты растений и животных. Близость организации энергетических систем у а-протеобактерий (таких как Rickettsia prowazekii) и митохондрий отражает существование у них общего предка и позволяет предполагать, что предок митохондрий представлял собой факультативный аэроб из семейства а-протеобактерий. Однако прокариотический предок митохондрий должен был обладать геномом, отличным от генома современных митохондрий, поскольку в результате их эволюции многие гены переходили в ядерный геном или просто терялись. Не все события эндосимбиоза, подобно митохондриям и хлоропластам, закрепились в процессе эволюции. Некоторые взаимоотношения между организмами оформились «по необходимости». Это относится к случаю эндосимбиоза между азотфиксирующими бактериями и их хозяевами — растениями. Усвоение атмосферного азота (процесс его превращения в аммиак) осуществляется только некоторыми прокариотами, хотя все живые организмы для своей жизнедеятельности нуждаются в аммиаке. Как показано на рисунке ниже, представители прокариотической группы Rhizobium способны заселять корневые пили некоторых растений, образуя колонии микробов, называемые клубеньками. Процесс инициации заселения корней и образования клубеньков происходит по сигналу, вырабатываемому Rhizobium (Nod факторы).
Эти факторы начинают экспрессироваться, когда Rhizobium обнаруживает флавоноиды, специфичные для каждого вида растений. Поэтому клубеньки образуются только в том случае, когда прокариоты обнаружат специфический флавоноид растения и начнут синтезировать факторы Nod. В свою очередь, эти факторы будут осуществлять эндосимбиоз только с тем растением, которое секретирует данный флавоноид. Факторы Nod представляют собой молекулы олигосахаридов. Сейчас ведется поиск рецепторов этих факторов в клетках корневой системы растений. При исследовании связывания на уровне клеточных экстрактов уже идентифицированы возможные кандидаты на роль рецепторов Nod. Впрочем, идентифицированные белки связываются с факторами неспецифическим образом и поэтому вряд ли представляют собой рецепторы Nod in vivo. Еще один подход к поиску рецепторов Nod заключается в идентификации растительных белков, способных связывать сахара, а затем в исследовании их сродства и связывания с факторами Nod. Этот подход был использован для идентификации белка, называющегося лектин-нуклеотид фосфогидролаза (LNP), который специфически связывается с факторами Nod. Антитела к этому белку блокируют образование клубеньков, что позволяет предполагать, что LNP является существенным компонентом их формирования. В настоящее время предпринимаются попытки идентифицировать другие рецепторы Nod.
Эндосимбиоз играет роль в развитии и обеспечении жизнеспособности многих насекомых. К числу наиболее хорошо известных случаев такого эндосимбиоза относятся взаимоотношения между гороховой тлей и бактериями, относящимися к виду Buchnera. Эти бактерии относятся к Proteobacterium и являются облигатными эндосимбионтами, обитающими в клетках хозяина — тли. Buchnera существует в форме бактериоцитов, представляющих собой специализированные клетки, находящиеся в полости тела тли.
Перенос бактерии в яйца и эмбрионы свидетельствует в пользу того, что Buchnera инфицирует каждое насекомое. Почему гороховая тля переносит вторжение в свой организм бактерий Buchnera?. Оказывается, прокариоты пополняют рацион тли, обеспечивая ее несколькими необходимыми питательными веществами. Основным продуктом питания тли служит сок растений, богатый углеводами и почти не содержащий аминокислот. Трудно доказать экспериментально, какими питательными веществами бактерии обеспечивают клетки организма хозяина. Известно, однако, что в геноме Buchnera присутствуют гены, кодирующие ферменты синтеза аминокислот, которые необходимы для жизнедеятельности тли. Более того, насекомые, инфицированнные Buchnera, выживают на синтетическом корме, в составе которого незаменимые аминокислоты отсутствуют. Однако, если насекомых обработать антибиотиками, которые уничтожают Buchnera, они начинают голодать и быстро погибают. Эти данные однозначно свидетельствуют о том, что Buchnera служат источником аминокислот для клеток организма хозяина. Интересно, что тли, в организме которых не содержится Buchera, плохо растут даже при скармливании им богатого рациона, содержащего все незаменимые аминокислоты. Это позволяет предполагать, что при эндосимбиозе Buchera обеспечивает организм хозяина не только незаменимыми аминокислотами.
Клубеньки, образующиеся при инфицировании корней некоторых растений бактериями Rhizobium.
Этот эндосимбиоз выгоден растению: бактерии синтезируют аммиак,
который используется растением в процессе фиксации атмосферного азота. Бактериоциты представляют собой клетки,
находящиеся в организме гороховой тли (насекомого), инфицированной бактериями Buchera.
На фотографии показан окрашенный препарат бактериоцита,
в котором видно ядро и бактерии, заполняющие цитоплазму.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Как составлялся геном эукариот: эндосимбиоз VS. непрерывный горизонтальный перенос
Новость
Автор
Редактор
Результаты работы крупной международной группы ученых, опубликованные в августовском Nature, добавили огоньку в давнее противостояние двух точек зрения на появление в эукариотическом геноме генов прокариот. Одна концепция утверждает, что прокариотические гены попали в ядерные клетки главным образом вместе с предками органоидов, другая — что горизонтальный перенос генов в мире эукариот — процесс распространенный и непрерывный. Новая работа подтачивает фундамент второй концепции, приводя альтернативные объяснения очевидной неоднородности распределения генов у эукариот.
Ядерный геном эукариот — это своеобразная мозаика из ДНК, приобретенной из различных источников. Например, из митохондрий и хлоропластов — эукариотических органоидов, которые произошли от прокариотических клеток, ассимилированных в процессе эндосимбиоза. Хотя эти органоиды и сохраняют часть своей первоначальной ДНК, внушительное количество их наследственного материала переместилось в ядро хозяина вследствие эндосимбиотического переноса генов [1]. В случае с хлоропластами дальнейшие эукариот-эукариотические эндосимбиозы распространили эти органоиды и связанные с ними гены по ветвям эволюционного дерева [2].
У прокариот вертикальное наследование (клональный рост) дополняется процессом рекомбинации в результате горизонтального переноса генов (ГПГ)*, осуществляемого путем трансформации, конъюгации, трансдукции. Эти механизмы работают однонаправленно — от донора к реципиенту — и приводят к формированию пангеномов [3].
* — О горизонтальном генетическом переносе у прокариот и многообразии его виновников — мобильных генетических элементов — рассказывает статья «Мобильные генетические элементы прокариот: стратификация „общества“ бродяжек и домоседов» [4].
Для эукариот характерна половая рекомбинация — процесс двунаправленный, обоюдный, так как перекраивается и отцовская, и материнская ДНК. Наследование же происходит почти исключительно вертикально из-за дефицита систем ГПГ, подобных прокариотическим. Отделение половых клеток от остальной части организма у многоклеточных эукариот, как полагают, служит прочным барьером для интродукции чужеродной ДНК. Однако это предположение многие исследователи считают сомнительным и приводят примеры горизонтального переноса генов и у животных, и у растений:
- вирусы могут служить векторами для переноса генов и облегчать ГПГ у эукариот так же, как у прокариот [5];
- одноклеточные эукариоты могут приобретать чужеродную ДНК, просто поглощая и переваривая добычу.
Никто не отрицает эпизод мощного вброса прокариотических генов при «одомашнивании» хлоропластов и митохондрий, когда часть наследственного материала эндосимбионтов переместилась в ядро клетки [6]. Однако большинство случаев горизонтального переноса среди эукариот было выявлено исключительно на основе филогенетических несоответствий — обнаружения генов, эволюционные истории которых не согласуются с известными или предсказанными взаимоотношениями организмов. Здесь следует учесть еще и то, что при построении деревьев часто возникают артефакты, отчего страдает воспроизводимость результатов*, разнятся их интерпретации [7].
* — Проблемы с воспроизводимостью и достоверностью результатов биологических, медицинских, психологических исследований довели-таки научное сообщество и спонсоров до отчаяния — до тайных собраний, отказа от традиционных критериев, массового освоения интернет-облаков и генерации прожектов на тему открытой и честной науки: «Мечты о воспроизводимости» [8].
Еще более спорное утверждение — об обильном и постоянном горизонтальном переносе генов от прокариот к эукариотам [9]. Если это верно, то следствием таких процессов должен быть кумулятивный эффект в эволюции эукариот, который должен обнаруживаться при анализе большого числа геномов разных эукариотических линий. Эта гипотеза противопоставляется теории эндосимбиоза, которая утверждает, что приобретение чужеродных генов эукариотами соответствует времени происхождения органоидов и вторичного эндосимбиоза у водорослей (рис. 1).
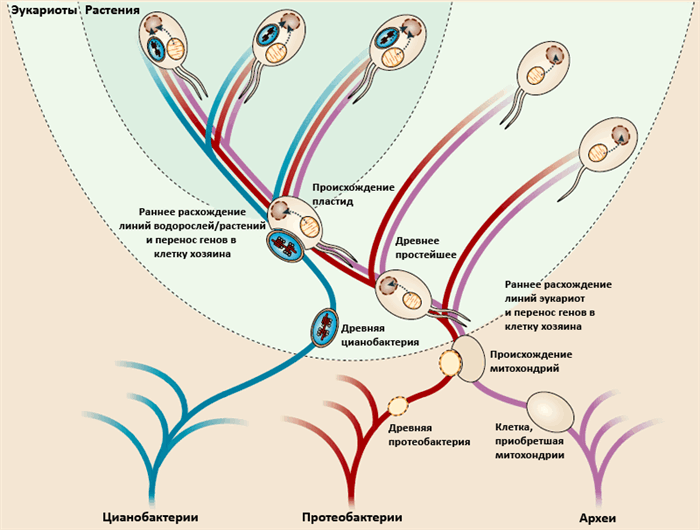
Рисунок 1. Эндосимбиотическая эволюция. Внутриклеточные эндосимбионты, которые первоначально произошли от свободноживущих прокариот, сыграли важную роль в эукариотической эволюции, породив два цитоплазматических органоида. Митохондрии развились из α-протеобактерий, а хлоропласты — из цианобактерий. Оба органоида внесли весомый вклад в совокупный генόм эукариотических ядер. На рисунке показана схема эволюции эукариот с акцентом на моментах включения митохондрий и хлоропластов в эукариотическую линию посредством эндосимбиоза и дальнейшей параллельной эволюции ядерного и органоидных геномов. Клетка-хозяин, которая приобрела пластиды, вероятно, обладала двумя жгутиками. Общепринято, что хозяин, заполучивший митохондрии (внизу справа), состоял в родстве с археями, однако о характере его внутриклеточной организации (прокариотической, эукариотической или промежуточной), возрасте, биохимии и генном составе биологи пока договориться не могут. Полагают, что хозяин был одноклеточным. Рисунок из [2].
Основание для заявлений о широком распространении горизонтального переноса генов у эукариот возникло после выявления неожиданных ветвей филогенетических деревьев и неоднородного распределения генов между линиями. Действительно, некоторые случаи линия-специфичного горизонтального переноса генов в эукариотические клетки неопровержимы: это включения бактериальных генов в хромосомы насекомых и вирусные вставки в процессе эволюции плацентарных [10, 11]. И всё же, если ГПГ у эукариот — непрерывный процесс, он должен привести к кумулятивному эффекту. Даже при очень низкой средней частоте переноса чужой ДНК в геном эукариот (например, 0,5%) приобретенные гены будут наследоваться потомками, которые продолжат приобретать новые гены. В итоге должны появляться всё более и более различающиеся генетические линии эукариот, которые будут со временем расходиться всё дальше друг от друга. Как раз такую схему эволюции мы наблюдаем в прокариотическом мире, где работает в полную силу ГПГ и накапливаются пангеномы.
Так как же всё-таки объяснить присутствие прокариотических генов в хромосомах эукариот — постепенным их накоплением в результате ГПГ или эпизодическим заимствованием от органоидов? Международная группа ученых из Германии, Новой Зеландии, Израиля, Португалии и других стран попыталась тщательно проверить обе гипотезы [3].
Распределение генов подтверждает теорию эндосимбиоза
В исследовании использовали около миллиона белковых последовательностей 55 эукариот шести супергрупп (рис. 2) и более шести миллионов последовательностей прокариот (300 000 от 134 архей, остальные — от 1847 бактерий). В результате выявили 2585 обособленных семейств белковых последовательностей, каждое из которых включало связанные (гомологичные) белки, кодируемые двумя или более эукариотическими и пятью или более прокариотическими геномами: эти семейства были названы эукариот-прокариотическими кластерами (ЭПК). Остальные — эукариотспецифичными кластерами (ЭСК).
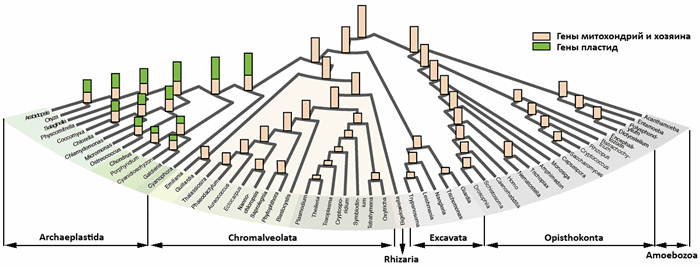
Рисунок 2. Эволюция эукариотического генома. Внизу показаны 6 супергрупп эукариот. Гены, унаследованные от эукариотического предка и митохондрий, обозначены пшеничным цветом, гены пластидного происхождения — зеленым (у организмов со вторичными пластидами не обозначены). Высота вертикальных прямоугольников отражает размер геномов (число ЭПК) «узловых» эукариот. Заметно, что эволюция эукариот сопровождалась потерей генов. Последовательности архейного происхождения (преимущественно «информационные»), унаследованные от линии архей, захватившей предка митохондрии и развившейся в эукариот, утрачивались реже, чем прочие гены («операционные», метаболические) [3]. Рисунок из [3], изменен. Нажмите на изображение, чтобы увеличить (откроется в отдельном окне).
87% деревьев первого кластера показали эукариотическую монофилию, для 12% она не исключалась. Однако остальные вовсе не обязательно представляют собой продукты горизонтального переноса генов, поскольку: а) эндосимбиоз привносит генетическую избыточность (например, в «хозяйскую» линию проникают гены рибосом эндосимбионтов); б) у этих деревьев выявлены признаки загрязнения при секвенировании; в) иногда молекулярная филогенетика. просто ошибается. Даже если предположить, что эти деревья отражают результат ГПГ, эукариоты всё равно не расширили свой генный репертуар за счет принятых последовательностей, поскольку повторно приобрели гены, уже присутствующие в эукариотической линии.
Сделав такой вывод, исследователи сфокусировались на изучении ЭПК-деревьев с эукариотической монофилией. Больше половины из этих кластеров (1060) имели прямое отношение к внедрению фотосинтеза в эукариотическую линию и его распространению путем вторичного эндосимбиоза (при формировании красных вторичных пластид страменопил, альвеолят, Guillardia, Emiliania и зеленых вторичных пластид Bigelowiella) [12, 13].
В той же группе (ЭПК) лишь единичные прокариотические кластеры были монофилетическими, что отражает ситуацию с пангеномами и распространенным среди прокариот горизонтальным переносом генов. Если бы эукариоты были вовлечены в пангеномный перенос генов вместе с прокариотами, они бы тоже обладали пангеномами, чего не наблюдается. Это наводит на мысль, что, за исключением эпизодов «приручения» предков митохондрий и пластид, эукариоты были изолированы от ГПГ, ведущего к формированию пангеномов у прокариот.
Эволюция эукариотических генов вертикальна
Существует только два механизма, которые могли привести к образованию 87% монофилетических деревьев в группе ЭПК:
- ген белка присутствовал в общем предке и вертикально наследовался потомками;
- ген был приобретен одним из членов группы и затем распространен посредством эукариот-эукариотического ГПГ.
В первом случае деревья на всём своём протяжении будут сходны с деревьями ЭСК. Во втором они будут отличны друг от друга и от каждого следующего вновь возникшего.
Ученые показали, что эволюция эукариотических генов «вопиюще» вертикальна, в то время как альтернативный механизм, связанный с эукариот-эукариотическим горизонтальным переносом генов, полностью исключили для всех шести супергрупп. В качестве доказательства они привели пример с супергруппой архепластид (растений в широком смысле). Анализ ЭПК-деревьев показал, что:
- хотя совсем немного архепластидспецифичных ЭПК можно отследить прямо до цианобактерий, все архепластидспецифичные ЭПК представлены у общего предка Archaeplastida;
- появление этих ЭПК совпадает с образованием пластид;
- многие кластеры имеют непосредственное отношение к фотосинтезу;
- но: их сестринские прокариотические группы филогенетически неоднородны, то есть гомологичные гены есть у разных прокариот (не только у цианобактерий).
Последнее наблюдение можно объяснить двумя способами.
- Если считать эти прокариотические таксоны биологическими донорами, то тогда в происхождение пластид должны быть вовлечены сотни независимых горизонтальных переносов генов от сотен различных доноров. То есть фотосинтезирующие эукариоты должны были строиться ген за геном, при этом исключались всякие мутации, инактивирующие перенесенные гены до сборки пластиды в готовую функциональную единицу.
- Деревья вводят в заблуждение, и архепластидспецифичные ЭПК приобретены от предка пластид, уже имеющего функционирующий фотосинтетический аппарат, который нужно было просто интегрировать в эукариотическую линию посредством переноса необходимых генов из органеллы в ядро (эндосимбиотического переноса генов).
Второй вариант, безусловно, более правдоподобен (рис. 3).
Эпизодический приток и избирательная потеря генов
О митохондриях можно рассуждать по аналогии с пластидами. Хозяин пластид изначально был гетеротрофом, а переход к автотрофии произошел посредством эндосимбиоза с переносом генов. Распределение генов, полученное в новом исследовании, это доказывает. Хозяевами митохондрий на каком-то этапе стали археи*, а эндосимбиоз с переносом генов из органеллы в ядро хозяина привел к возникновению хемиосмотического синтеза АТФ. Как и пластиды, митохондрии не могли быть сконструированы пошагово, путем последовательных горизонтальных переносов генов: сотни беспорядочно приобретенных для создания дыхательного органоида генов не могли поддерживаться в клетке очищающим отбором до момента сборки полноценной митохондрии. Перенести необходимые гены в ядро из дышащего эндосимбионта было бы несравнимо проще (рис. 3).
* — Особенности эволюции архей, насыщенной судьбоносными заимствованиями генов у бактерий, подробно разобраны в статье «Закинули археи эволюционный невод и вытянули. » [14].
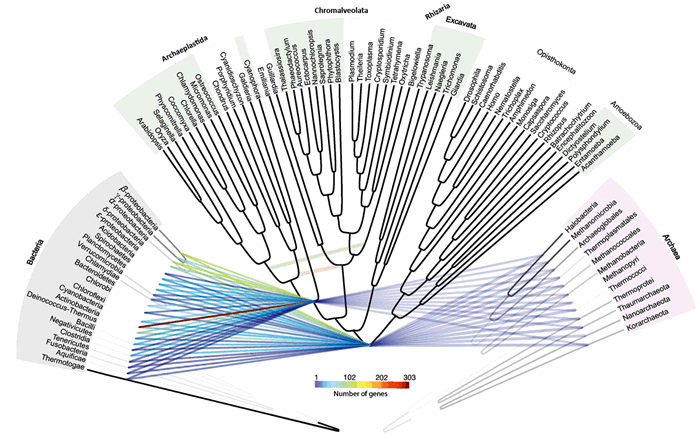
Рисунок 3. Сеть эндосимбиотического переноса генов (ЭПГ). На карту референсной филогении нанесена сеть, соединяющая предполагаемых доноров генов с общими предками архепластид и всех эукариот. Интенсивность серых линий (от белого до черного) в прокариотических референсных деревьях показывает, насколько часто конкретная ветвь «перекрывалась» дендрограммами отдельных генов, использованных для формирования итоговой топологии. Цветные линии показывают приток генов в точках происхождения эукариот и образования пластид. Цвет линий (по шкале) отражает, сколько общих генов обнаружено у эукариот с тем или иным прокариотическим таксоном (т.е. возможный донорский вклад конкретной группы бактерий или архей). Вторичные ЭПГ обозначены изолированными молочно-зелёной и молочно-красной линиями. Рисунок из [3]. Нажмите на изображение, чтобы увеличить (откроется в отдельном окне).
Но как объяснить неоднородное распределение генов между линиями эукариот, о котором говорилось в начале статьи? У прокариот такое положение дел указывает на ГПГ (за исключением изолированных видов с редукционной эволюцией), что подталкивает к подобному выводу и в отношении эукариот. Однако, как показали ученые, вертикальное наследование у эукариот характеризуется дифференциальной (избирательной) потерей генов, которая наилучшим образом объясняет неоднородное распределение генетической информации в этом домене. Это справедливо как для ЭПК, так и для ЭСК. В то же время приобретения новых генов — события эпизодические, соответствующие периоду формирования эндосимбионтов.
Некоторые наборы генов из эукариот-прокариотических кластеров кажутся специфичными для конкретных линий приобретениями: например, 67 генов у диктиостелид, 160 генов у опистоконтов и 210 генов, предположительно приобретенных предком сухопутных растений. Если эти гены были заимствованы путем горизонтального переноса недавно, например, в период становления сухопутных растений 450 млн лет назад, они должны быть более сходными с их прокариотическими «родственниками», чем гены, полученные от эндосимбионтов. Однако обнаружено обратное. Гены-кандидаты, подозреваемые в геном-специфичном (горизонтальном) проникновении из про- в эукариот, тоже были выявлены, но встречаются эти предполагаемые продукты ГПГ приблизительно в 4-10 раз реже, чем ядерные вставки митохондриальных и пластидных последовательностей, а число их уменьшается по мере обновления описаний геномов.
Заключение
Анализ распределения эукариот-прокариотических кластеров выявил ожидаемо большой эволюционный след, оставленный приобретением хлоропластов. Сотни ядерных генов отсутствуют у эукариотических видов, не имеющих хлоропластов, но распространены среди различных групп эукариот-фотосинтетиков и цианобактерий, от которых хлоропласты и произошли. Эндосимбиотический след в митохондриальной эволюции тоже очевиден, хотя менее отчетлив.
- перенесенные горизонтально гены, специфичные для определенных линий, быстро элиминируются;
- восприимчивые к ГПГ линии не дают начало эволюционно стабильным потомкам (тупиковы);
- многие предполагаемые продукты ГПГ на самом деле не являются линия-специфичными, и при расширении аналитической выборки эукариот они будут обнаружены у других отдаленных родственников, что докажет причинную роль дифференциальной потери в возникновении неоднородности распределения генов;
- возможна любая комбинация описанных вариантов.
Итак, по мнению участников исследования [3], два эпизода эндосимбиотического переноса генов (один — от митохондрий, другой — от пластид) с последующей избирательной потерей ДНК объясняют филогению и неоднородное распределение бактериальных генов в эукариотах. Кроме того, авторы дали возможность оценить достоинства и недостатки разных подходов, применяемых для различения горизонтального и вертикального наследований у эукариот. Но, без сомнения, вопрос остается открытым для обсуждения.
Эндосимбиоз: теория в состоянии кризиса
Одна из величайших проблем теории эволюции заключается в невозможности объяснить, как клетки растений и животных, эукариоты, могли произойти от своих предшественников, бактериеобразных клеток, называемых прокариотами. В отличие от прокариотов, клетки эукариотов состоят из нескольких отделов и содержат заключенные в мембраны органоиды, такие как ядро, митохондрия, и (в растениях) хлоропласты, которых нет в бактериях и археях (небактериальных одноклеточных прокариотах). Наряду со многими другими генетическими и молекулярными различиями, эти сложные клеточные органоиды, отсутствующие в прокариотах, представляют собой непреодолимую для теории эволюции пропасть. 1, 2
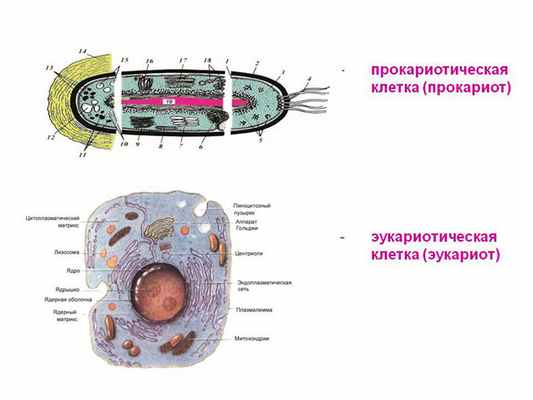
Главным различием прокариот и эукариот является то, что у прокариот нет оформленного ядра, отделенного мембраной от цитоплазмы. У прокариот кольцевая ДНК находится в цитоплазме, а место, где находится ДНК, называется нуклеоидом.
Также, из органоидов прокариоты имеют только рибосомы 70S (мелкие), а у эукариот имеются не только крупные 80S рибосомы, но и много других органоидов.
Так как ядра у прокариот нет, то делятся они делением надвое – не с помощью мейоза/митоза.
Эукариоты имеют гистоны, которых нет у бактерий. Хромантин эукариот содержит 1/3 ДНК и 2/3 белка, у прокариот все наоборот.
Клетка эукариот в 1000 раз больше по объему и в 10 раз больше по диаметру, чем клетка прокариот.
Эволюционисты уже давно пытаются представить неспекулятивное объяснение клеточного ядра эукариота, а также других сложных особенностей клетки. На протяжении почти 50 лет было довольно популярным представление о том, что митохондрии и хлоропласты появились в результате некоего мифического процесса под названием эндосимбиоз. 3 В основе этого объяснения лежит тот факт, что митохондрион обладает небольшим кругообразным отрезком ДНК, в котором содержатся гены, необходимые ему для функционирования; в нем также предусмотрен процесс репродукции органоидов. Эта функциональная комбинация напоминает по своему строению и особенностям репродукции бактерии, но это сходство поверхностное.
Предполагается, что в какой-то момент в отдаленном прошлом некая прокариотическая клетка (возможно, архея) поглотила бактерию (возможно, протеобактерию), а затем, в ходе некоего магического скачка, заменяющего любое научное объяснение, сотни генов были каким-то образом «перекроены» для выполнения новых функций. Кроме того, считается, что многие тысячи генов были перемещены в клеточные ядра, тогда как другие были отбракованы. Базовые идеи всего этого сценария были выдвинуты в результате обнаружения генов как в геноме митохондрий, так и в геноме эукариотов, в которых некоторые участки имеют сходство с генами бактерий (в особенности протеобактерий). 3
Однако на сегодняшний день, когда определение последовательности генома стало широко распространенным и не слишком дорогостоящим процессом, эволюционная история об эндосимбиозе становится все более туманной и противоречивой. По мере того, как устанавливается все больше последовательностей генома бактерий и эукариотов и определяется, какие белки они кодируют, вся эволюционная идея эндосимбиоза становится все более запутанной.
Одним из самых неожиданных открытий стало отсутствие генов, способных подтвердить эту эволюционную сказку. В одной из недавно вышедших статей отмечается:
«Чего мы не предвидели – так это того, что относительно небольшое количество митохондриальных белков с бактериальной гомологией [сходством последовательности] будут особым образом группироваться с α-протеобактерией в филогенетических реконструкциях [реконструкциях эволюционного дерева]: в лучшем случае, лишь 10–20% всех исследованных на данный момент белковых карт демонстрируют активную α-протеобактериальную связь». 4
Отсутствие доказательств того, что митохондриальные гены имеют бактериальное происхождение как в митохондриальной ДНК, так и в геноме ядра, где содержится большинство митохондриальных генов, представляет собой серьезную проблему для эволюционной истории.
Еще одно важное открытие, противоречащее эволюционным предположениям об эндосимбиозе, заключается в отсутствии картины устойчивой эволюционной прогрессии от простого к сложному. Митохондриальные белковые карты (весь набор митохондриальных белков) даже для таких одноклеточных организмов, как грибки и простейшие, уже невероятно сложны, многообразны и полноценны. Кроме того, наборы митохондриальных генов одноклеточных эукариотов так же сложны, как и гены растений и животных. Как отметил один из ведущих ученых-эволюционистов, «становится очевидным, что «последний митохондриальный общий предок» был изначально таким же сложным по своим ключевым функциям, как и современная митохондрия» и «очевидно, базовый «перекрой» от эндосимбионта к митохондриону был, фактически, завершен, до начала процесса дивергенции на основные генеалогические линии эукариотов» . 4
Последовательности геномов множества эукариотов четко указывают на то, что набор генов, необходимый митохондрии для функционирования, не произошли от бактерии, а уникальны для того вида живых существ, в которых они обнаруживаются. Хотя определенные генетические сходства действительно существуют, эти сходства убедительно объясняются стандартной инженерной концепцией повторного использования кодов (когда некий общий код решает схожие проблемы). Богатство геномных данных на сегодняшний день в корне разрушает идею эволюции на всех фронтах, даже в сфере эндосимбиоза – одной из излюбленных теорий секуляристов.
Ссылки и примечания
- Тэн К. и Д. Томкинз s. 2015. Различия в обработке информации у архей и эукаритов. Значение для гомологов и для мифа об эукариогенезе. Издание Answers Research Journal. 8: 121–141. Вернуться к тексту.
- Тэн К. и Д. Томкинз. 2015 Различия в обработке информации у архей и эукаритов. Значение для гомологов и для мифа об эукариогенезе. Издание Answers Research Journal.8: 143–162. Вернуться к тексту.
- Мартин В. и др.. 2001. Обзор эндосимбиотических моделей происхождения эукариотов, их АТП-образующие органоиды (митохондрии и гидрогеносомы), а также их гетеротрофный образ жизни. Издание. Biological Chemisty. 382 (11): 1521–1539. Вернуться к тексту.
- Грей М.В. 2015. Мозаичная природа митохондриального протеома: выводы относительно происхождения и эволюции митохондрии .Издание. Proceedings of the National Academy of Sciences. 112 (33): 10133–10138. Вернуться к тексту.
* Доктор Томкинз – научный сотрудник Института исследования сотворения; он получил степень кандидата наук в сфере генетики в Университете Клемсон.
Эндосимбиоз прокариот и эукариот

Предки всех эукариот
В океане, на глубине 3 283 метра, между Гренландией и Норвегией учёные обнаружили фрагменты ДНК неизвестного организма. Когда изучили геном, выяснили, что это тот самый загадочный родоначальник всех живых существ с клеточным ядром. Всех многоклеточных животных и растений, а также амёб, прочих инфузорий и нас с вами.
Прокариоты — одноклеточные организмы, устроенные намного проще, чем эукариоты. Прокариоты — бактерии и археи. У них нет ядра и органелл, то есть всё содержимое клетки плавает в цитоплазме. Их ДНК устроена просто: она не делится на хромосомы (чаще всего ДНК замкнута в колечко).
Археи — одноклеточные организмы, отличающиеся и от бактерий, и от эукариот. Один из трёх главных доменов (высшая категория в систематике живых организмов). Не имеют ядра и мембранных органелл. Считаются самыми древними по происхождению из ныне живущих существ.
Клеточные мембраны — двойной слой липидных молекул с включёнными в него молекулами белков. Отделяют клетку от окружающей среды, ядро клетки — от цитоплазмы, клеточные органеллы — от других органелл. Кроме того, выполняют роль энергетических станций — в митохондриях и хлоропластах. В целом клеточные мембраны — главное, что позволило организмам стать многоклеточными.[/note][/pullquote]
«Первые эукариоты произошли от архей» — к такому выводу пришли учёные из Упсальского университета (старейший университет Швеции). Тийс Эттема с коллегами исследовали фрагменты ДНК, найденные в донных отложениях гидротермального источника Замок Локи. Фрагменты были намного короче всей ДНК организмов, которым они принадлежали, но поскольку кусочков было много и они перекрывались между собой, по ним удалось восстановить структуру ДНК их хозяев. Всего собрали три генома, один получился почти полным — получилось аккуратно восстановить 92% его ДНК. Обладатель этого генома был археем — простым одноклеточным организмом, ДНК которого не заключена в ядро, а свободно плавает в цитоплазме. Организмы получили название Lokiarcheum, сокращённо — Локи.
Эукариоты произошли от прокариот — в этом нет сомнений. Произошли очень давно, 1,5–2 млрд лет назад, на заре эволюции. Но как это получилось, откуда вдруг в одноклеточных организмах, похожих на примитивные бактерии, взялось ядро и множество других сложных структур? До сих пор это было не очень понятно. Предполагалось, что вся сложность от архей — поныне существующих одноклеточных, почти таких же, как бактерии, но очень древних, судя по строению молекулярных комплексов. Экспериментальных подтверждений этой гипотезе было совсем мало, к тому же оставался вопрос: есть ли общий предок у архей и эукариот, или когда-то археи сами мутировали и произвели клеточное ядро на нашу радость?
Образование ядра — ключевое событие всей эволюции. Наравне с ещё двумя: собственно происхождением жизни и появлением многоклеточных существ. Все три события — тайна, покрытая мраком миллиардов лет. Мы наблюдаем в реальном времени, как раскрывают одну из этих тайн. Захватывающе.
Итак, расшифровали ДНК организмов, поднятых со дна моря. Что дальше?
Большинство генов Локи оказались архейными, то есть прокариотическими. Но обнаружилось среди них и несколько генов, важных для эукариотических клеток и поддержания сложной жизни их внутренних мембран. Во-первых, это были гены актина — белка, который составляет внутренний скелет эукариотических клеток и заодно может служить «рельсами», по которым транспортируются мембранные пузырьки с запасами полезных молекул. Ещё актин необходим для фагоцитоза — захвата чужих клеток и их переваривания. У людей, например, это оружие иммунитета: фагоциты захватывают бактерии и уничтожают их. А два миллиарда лет назад такого не было, но актин мог пригодиться на следующей ступеньки эволюции: большие одноклеточные захватили навсегда бактерий, интегрировали их (буквально) и сделали из них митохондрии — важнейшую часть всех клеток, которая производит энергию.
Локи перевернул представления учёных о развитии жизни, потому что раньше сложные манипуляции с мембранами считались прерогативой эукариот. Теперь получается, что ещё до поглощения внутриклеточных симбионтов — митохондрий, с которыми возникли эукариоты, — их предок уже обладал начальными способностями к обращению с мембранами, а потому был готов принять внутренних симбионтов. И этот предок эукариот, как и Локи, был археем.
Древо жизни в представлении учёных изменило свой облик: раньше бактерии, археи и эукариоты представляли на нём три отдельные ветви, а теперь ветвь эукариот отходит от архей. Оказалось, что они не так уж и непохожи на эукариот и могут заполнить пропасть между ними и проще устроенными прокариотами.
Конечно, теперь научное сообщество с нетерпением ждёт обнаружения живых хозяев восстановленного генома. Искать Локи, скорее всего, станут там, где будет выявляться больше их специфической ДНК.
Комментарий эксперта
Евгений Кунин — ведущий научный сотрудник Национального центра биотехнологической информации (National Center for Biotechnology Information, Бетесда, США), эксперт с мировым именем в области компьютерной и эволюционной биологии:
«Bridge the gap (заполнили разрыв. — КШ) — это, конечно, метафора и, как почти любая метафора, неточность. Однако открытие очень важное, и выводы в самой статье вполне корректны. Описанный организм, насколько можно судить по геному, действительно представляет собой архею со сложной внутриклеточной организацией и беспрецедентным набором эукариотических черт.
Речь идёт о сравнительно небольшом числе генов (хотя как считать — в этот набор входит очень даже солидное семейство регуляторных ГТФаз), но здесь важно не количество, а качество. Именно белки, кодируемые этими генами, обеспечивают формирование цитоскелета, а возможно, и внутриклеточных мембран. Эти внутриклеточные структуры вполне могут обеспечить возможность поглощения таким организмом других микробов, в частности бактерий, что создаёт предпосылки для эндосимбиоза. Разумеется, это всё гипотезы — экспериментальные данные могут появиться, только когда организм будет исследован в лаборатории, что крайне нелегко. Однако анализ генома очень чётко указывает на эти возможности. Замечу, что характеристики архейного предка эукариот были предсказаны — прямо скажу, в первую очередь в работах моей группы — на основе анализа геномов других архей. Точность, с которой Локи соответствует этим предсказаниям, впечатляет.
Крайне важен и другой аспект этой работы. Однозначно показано, что Локи — ближайший родственник эукариот среди известных архей. Эти результаты не оставляют сомнений в том, что эукариоты происходят, так сказать, из гущи архейного разнообразия. Таким образом, предположение, что эукариоты происходят от общего предка с археями, а не именно от определённой группы архей, не просто создаёт ненужную новую сущность — оно неверно. Открытие Локи, можно сказать, покончило с этой гипотезой.
Именно сочетание этих двух аспектов — открытие ближайшего родственника эукариот среди архей и демонстрация сложной внутриклеточной организации этого организма — делает открытие Локи настоящим прорывом в изучении происхождения эукариот. Bridge or no bridge, так сказать».
Читайте также: