Мембранный потенциал покоя. Потенциал покоя нервных клеток
Добавил пользователь Skiper Обновлено: 23.01.2026
Мембрана всех живых клеток поляризована. Внутренняя сторона мембраны несет отрицательный заряд по сравнению с межклеточным пространством (рис. 1). Величина заряда, который несет мембрана называется мембранным потенциалом (МП). В невозбудимых тканях МП низкий, и составляет около -40 мВ. В возбудимых тканях он высокий, около -60 - -100 мВ и называется потенциалом покоя (ПП).
Потенциал покоя, как и любой мембранный потенциал формируется за счет избирательной проницаемости клеточной мембраны. Как известно, плазмолемма состоит из липидного бислоя, через который движение заряженных молекул затруднено. Белки, встроенные в мембрану, могут избирательно изменять проницаемость мембраны для различных ионов, в зависимости от приходящих стимулов. При этом, для формирования потенциала покоя ведущую роль играют ионы калия, кроме них важны ионы натрия и хлора.
Рис. 1. Концентрации и распределение ионов с внутренней и внешней стороны мембраны.
Большинство ионов распределяются неравномерно с внутренней и внешней стороны клетки (рис. 1). Внутри клетки концентрация ионов калия выше, а натрия и хлора – ниже, чем снаружи. В состоянии покоя мембрана проницаема для ионов калия и практически непроницаема для ионов натрия и хлора. Несмотря на то, что калий может свободно выходить из клетки, его концентрации остаются неизменными благодаря отрицательному заряду на внутренней стороне мембраны. Таким образом, на калий действуют две силы, находящиеся в равновесии: осмотические (градиент концентрации К + ) и электрические (заряд мембраны), благодаря чему число входящих в клетку ионов калия равно выходящим. Движение калия осуществляется через калиевые каналы утечки, открытые в состоянии покоя. Величину заряда мембраны, при которой ионы калия находятся в равновесии можно вычислить по уравнению Нернста:
где Ек — равновесный потенциал для К + ; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; n — валентность К + (+1), [К + н] — [К + вн] — наружная и внутренняя концентрации К + .
Если подставить в уравнение значения из таблицы на рис. 43, то мы получим величину равновесного потенциала, равную примерно -95 мВ. Это значение вписывается в диапазон мембранного потенциала возбудимых клеток. Отличия ПП разных клеток (даже возбудимых) могут возникать по трем причинам:
- отличия внутриклеточной и внеклеточной концентраций ионов калия в разных тканях (в таблице приведены данные по среднестатистическому нейрону);
- натрий-калиевая АТФаза может вносить свой вклад в значение заряда, так как она выводит из клетки 3 Na + в обмен на 2 К + ;
- несмотря на минимальную проницаемость мембраны для натрия и хлора, эти ионы все-таки могут попадать в клетки, хоть и от 10 до 100 раз хуже, по сравнению с калием.
Чтобы учесть проникновение других ионов в клетку существует уравнение Нернста-Гольдмана:
, где Еm — мембранный потенциал; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; РK , PNa и РCl — константы проницаемости мембраны для К + Na + и Сl, соответственно; [К + н], [K + вн], [Na + н], [Na + вн], [Сl — н] и [Сl — вн ]- концентрации K + , Na + и Сl снаружи (н) и внутри (вн) клетки.
Такое уравнение позволяет установить более точную величину ПП. Обычно, мембрана оказывается на несколько мВ менее поляризована, по сравнению с равновесным потенциалом для К + .
Потенциал действия (ПД) может возникать в возбудимых клетках. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то ПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет кратковременная перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной, после чего восстановится ПП. Это кратковременное изменение ПП, происходящее при возбуждении клетки называется потенциалом действия.
Возникновение ПД возможно благодаря тому, что в отличие от ионов калия, ионы натрия далеки от равновесия. Если подставить в уравнение Нернста натрий вместо калия, то мы получим равновесный потенциал, равный примерно +60 мВ. Во время ПД, происходит кратковременное увеличение проницаемости для Na + . При этом, натрий начнет проникать в клетку под действием двух сил: по градиенту концентрации и по заряду мембраны, стремясь подстроить заряд мембраны под свой равновесный потенциал. Движение натрия осуществляется по потенциал-зависимым натриевым каналам, которые открываются в ответ на смещение мембранного потенциала, после чего сами инактивируются.
Рис. 2. Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
На записи ПД выглядит как кратковременный пик (рис. 44), имеющий несколько фаз.
- Деполяризация (фаза нарастания) (рис. 44) – увеличение проницаемости для натрия из-за открытия натриевых каналов. Натрий стремится к своему равновесному потенциалу, но не достигает его, так как канал успевает инактивироваться.
- Реполяризация – возвращение заряда к величине потенциала покоя. Помимо калиевых каналов утечки здесь подключаются потенциал-зависимые калиевые каналы (активируются от деполяризации). В это время калий выходит из клетки, возвращаясь к своему равновесному потенциалу.
- Гиперполяризация (не всегда) – возникает в случаях, если равновесный потенциал по калию превышает по модулю ПП. Возвращение к ПП происходит после возвращения к равновесному потенциалу по К + .
Во время ПД происходит изменение полярности заряда мембраны. Фаза ПД, при которой заряд мембраны положителен, называется овершутом (рис. 2).
Благодаря Н-воротам инактивация канала происходит раньше, чем потенциал на мембране достигнет равновесной величины по натрию. После прекращения поступления натрия в клетку, происходит реполяризация за счет выходящих из клетки ионов калия. При этом к каналам утечки в этом случае подключаются еще и потениал-активируемые калиевые каналы. Во время реполяризации, в быстром натриевом канале быстро закрываются М-ворота. Н-ворота открываются гораздо медленнее и остаются закрытыми еще некоторое время после возвращения заряда к потенциалу покоя. Этот период принято называть периодом рефрактерности.
Рис. 3. Работа потенциал-управляемого натриевого канала.
Концентрации ионов внутри клетки восстанавливает натрий-калиевая АТФаза, которая с затратой энергии в виде АТФ откачивает из клетки 3 иона натрия и закачивает 2 иона калия.
По немиелинизированному волокну или по мембране мышцы потенциал действия распространяется непрерывно. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до порогового значения, в результате чего на соседнем участке возникает деполяризация. Главную роль в возникновении потенциала на новом участке мембраны предыдущий участок. При этом на каждом участки сразу после ПД наступает период рефрактерности, за счет которое ПД распространяется однонаправленно. При прочих равных условиях распространение потенциала действия по немиелинизированному аксону происходит тем быстрее, чем больше диаметр волокна. У млекопитающих скорость составляет 1-4 м/с. Поскольку у беспозвоночных животных отсутствует миелин, в гигантских аксонах кальмара скорость ПД может достигать 100 м/c.
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах немиелинизированных волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до порогового значения, что приводит к возникновению в них новых потенциалов действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-й, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Сальтаторное проведение увеличивает скорость проведения ПД 15-20 раз до 120 м/с.
Работа нейронов
Нервная система состоит из нейронов и глиальных клеток. Однако, главную роль в проведении и передаче нервных импульсов играют нейроны. Они получают информацию от множества клеток по дендритам, анализируют ее и передают или не передают на следующий нейрон.
Передача нервного импульса с одной клетки на другую осуществляется с помощью синапсов. Различают два основных типа синапсов: электрические и химические (рис. 4). Задача любого синапса – передать информацию с пресинаптической мембраны (мембрана аксона) на постсинаптическую (мембрана дендрита, другого аксона, мышцы или другого органа-мишени). Большинство синапсов нервной системы образуется между окончанием аксонов и дендритами, которые в области синапса образуют дендритные шипики.
Преимущество электрического синапса состоит в том, что сигнал с одной клетки на другую переходит без задержки. Кроме того, такие синапсы не утомляются. Для этого пре- и постсинаптические мембраны соединены поперечными мостиками, через которые ионы из одной клетки могут перемещаться в другую. Однако, существенным минусом такой системы является отсутствие однонаправленной передачи ПД. То есть, он может передаваться как с пресинаптической мембраны на постсинаптическую, так и наоборот. Поэтому, такая конструкция встречается достаточно редко и в основном – в нервной системе беспозвоночных.
Рис. 4. Схема строения химического и электрического синапсов.
Химический синапс весьма распространен в природе. О устроен сложнее, так как необходима система преобразования электрического импульса в химический сигнал, затем, вновь в электрический импульс. Все это приводит к возникновению синаптической задержки, которая может составить 0,2-0,4 мс. Кроме того, может произойти истощение запасов химического вещества, что приведет к утомлению синапса. Однако, такой синапс обеспечивает однонаправленность передачи ПД, что является его главным преимуществом.
Рис. 5. Схема работы (а) и электронная микрофотография (б) химического синапса.
В состоянии покоя окончание аксона, или пресинаптическое окончание, содержит мембранные пузырьки (везикулы) с нейромедиатором. Поверхность везикул заряжена отрицательно, чтобы предотвратить связывание с мембраной, и покрыта специальными белками, и принимающими участие в высвобождении везикул. В каждом пузырьке находится одинаковое количество химического вещества, которое называется квантом нейромедиатора. Нейромедиаторы весьма разнообразны по химическому строению, однако, большинство из них производятся прямо в окончании. Поэтому, в нем могут находиться системы, для синтеза химического посредника, а также аппарат Гольджи и митохондрии.
Постсинаптическая мембрана содержит рецепторы к нейромедиатору. Рецепторы могут быть в виде как ионных каналов, открывающихся при контакте со своим лигандом (ионотропные), так и мембранными белками, запускающими внутриклеточный каскад реакций (метаботропные). Один нейромедиатор может иметь несколько как ионотропных, так и метаботропных рецепторов. При этом, часть из них может быть возбуждающими, а часть – тормозными. Таким образом, реакцию клетки на нейромедиатор будет определять тип рецептора на ее мембране, и разные клетки могут совершенно по-разному реагировать на одно и то же химическое вещество.
Между пре- и постсинаптической мембраной располагается синаптическая щель, шириной 10-15 нм.
При приходе ПД на пресинаптическое окончание, на нем открываются потенциал-активируемые кальциевые каналы и ионы кальция входят в клетку. Кальций связывается с белками на поверхности везикул, что приводит к их транспортировке к пресинаптической мембране с последующим слиянием мембран. После такого взаимодействия нейромедиатор оказывается в синаптической щели (рис. 5) и может связаться со своим рецептором.
Ионотропные рецепторы – это лиганд-активируемые ионные каналы. Это значит, что канал открывается только в присутствии определенного химического вещества. Для разных нейромедиаторов это могут быть натриевые, кальциевые или хлорные каналы. Ток натрия и кальция вызывает деполяризацию мембраны, поэтому такие рецепторы называют возбуждающими. Хлорный ток приводит к гиперполяризации, что затрудняет генерацию ПД. Следовательно, такие рецепторы называют тормозными.
Метаботропные рецепторы к нейромедиаторам относят к классу рецепторов, ассоцированных с G-белками (GPCR). Эти белки запускают разнообразные внутриклеточные каскады реакций, приводящих в конечном итоге либо к дальнейшей передачи возбуждения, либо к торможению.
После передачи сигнала необходимо быстро удалить нейромедиатор из синаптической щели. Для этого в щели присутствуют либо ферменты расщепляющие, нейромедиатор, либо на пресинаптическом окончании или соседних глиальных клетках могут располагаться транспортеры, закачивающие медиатор в клетки. В последнем случае он может использоваться повторно.
Каждый нейрон получает импульсы от 100 до 100 000 синапсов. Одиночная деполяризация на одном дендрите не приведет к дальнейшей передаче сигнала. На нейрон могут приходит одновременно множество как возбуждающих, так и тормозных стимулов. Все они суммируются на соме нейрона. Такая суммация называется пространственной. Далее, может возникнуть или не возникнуть (в зависимости от пришедших сигналов) ПД в области аксонного холмика. Аксонный холмик – это область аксона, примыкающая к соме и обладающая минимальным порогом ПД. Далее импульс распространяется по аксону, конец которого может сильно ветвиться и образовывать синапсы со множеством клеток. Помимо пространственной, существует временная суммация. Она происходит в случае, поступления часто повторяющихся импульсов от одного дендрита.
Помимо классических синапсов между аксонами и дендритами или их шипиками, существуют также синапсы, модулирующие передачу в других синапсах (рис. 6). К ним относят аксо-аксональные синапсы. Такие синапсы способны усиливать или тормозить синаптическую передачу. То есть, если на окончание аксона, образующего аксо-шипиковый синапс, пришел ПД, а в это время по аксо-аксональному синапсу на него пришел тормозный сигнал, высвобождения нейромедиатора в аксо-шипиковом синапсе не произойдет. Аксо-дендритные синапсы могут изменять проведение мембраной ПД на пути от шипика к соме клетки. Также существуют аксо-соматические синапсы, которые могут влиять на суммацию сигнала в области сомы нейрона.
Таким образом, существует огромное многообразие различных синапсов, отличающихся по составу нейромедиаторов, рецепторов и их местоположению. Все это обеспечивает разнообразие реакций и пластичность нервной системы.
Мембранный потенциал покоя. Потенциал покоя нервных клеток
Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Плазматическая мембрана всех клеток, в том числе и нейронов, представлена двойным слоем (бислоем) фосфолипидов, в котором «головки» фосфолипидов обращены к водной среде внеклеточных и внутриклеточных пространств, а парные «хвосты» погружены внутрь и образуют липидную мембрану. Фосфатный слой растворим в воде (гидрофильный, или полярный), а двойной липидный слой — нерастворим (гидрофобный, или неполярный).
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl - ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl - ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl - на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
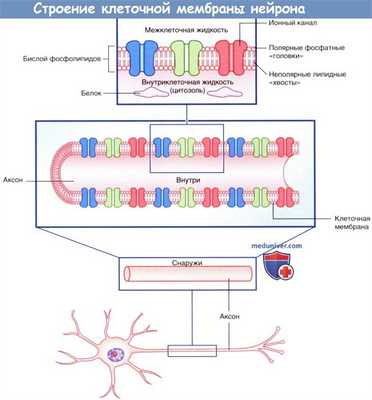
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl - . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl - намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
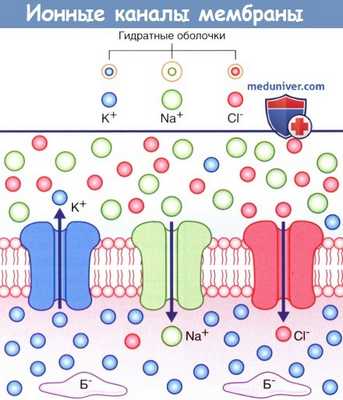
В состоянии покоя ионы Na + и Cl - сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б - ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl - во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl - — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
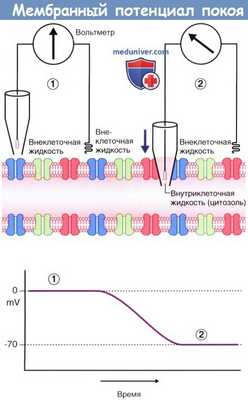
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl - :
Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl - внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
Уравнение Гольдмана сходно с уравнением Нернста, однако в нем перемножаются проницаемость мембраны для каждого иона и его концентрация; это уравнение применяют для установления потенциала «реверсии» через плазматическую мембрану с учетом ее проницаемости для разных ионов. Влияние ионов С1" на потенциал покоя имеет второстепенное значение, поскольку равновесный потенциал и потенциал покоя ионов Cl - приблизительно равны. Значение потенциала покоя в среднем составляет -70 мВ, и обеспечивается за счет суммарного влияния ионов К + и Na + .
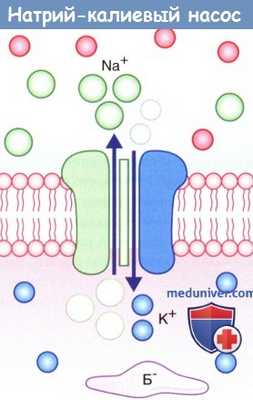
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б - — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.
Мембранный потенциал покоя крупных нервных волокон, когда по ним не проводятся нервные сигналы, составляет около -90 мВ. Это значит, что потенциал внутри волокна на 90 мВ отрицательнее, чем потенциал внеклеточной жидкости снаружи волокна. Далее мы объясним все факторы, определяющие уровень этого потенциала покоя, но прежде необходимо описать транспортные свойства мембраны нервного волокна для ионов натрия и калия в условиях покоя.
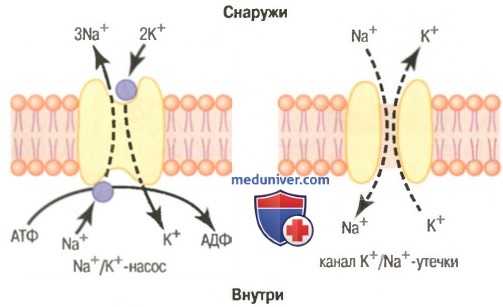
Функциональные характеристики Na+/K + -насоса и канала К + /Na + -утечки.
Активный транспорт ионов натрия и калия через мембрану. Натрий-калиевый насос. Вспомним, что все клеточные мембраны организма имеют мощный Na+/K+-Hacoc, постоянно выкачивающий ионы натрия наружу клетки и закачивающий внутрь нее ионы калия. Это электрогенный насос, поскольку положительных зарядов наружу перекачивается больше, чем внутрь (3 иона натрия на каждые 2 иона калия, соответственно). В результате внутри клетки создается общий дефицит положительных ионов, ведущий к отрицательному потенциалу с внутренней стороны клеточной мембраны. Na+/K+-Hacoc создает также большой градиент концентрации для натрия и калия через мембрану нервного волокна в покое:
Na + (снаружи): 142 мэкв/л
Na + (внутри): 14 мэкв/л
К + (снаружи): 4 мэкв/л
К + (внутри): 140 мэкв/л
Соответственно, отношение концентраций двух ионов внутри и снаружи составляет:
Na + внутри / Na + снаружи = 0,1
К + внутри / К + снаружи = 35,0
Утечка калия и натрия через мембрану нервного волокна. На рисунке показан канальный белок в мембране нервного волокна, называемый каналом калий-натриевой утечки, через который могут проходить ионы калия и натрия. Особенно существенна утечка калия, поскольку каналы более проницаемы для ионов калия, чем натрия (в норме примерно в 100 раз). Как обсуждается далее, это различие в проницаемости чрезвычайно важно для определения уровня нормального мембранного потенциала покоя.
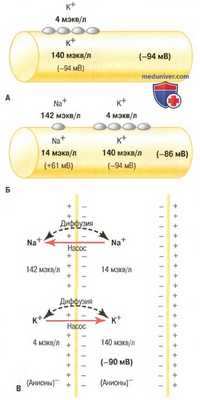
Формирование мембранных потенциалов в нервных волокнах при трех условиях.
А. Мембранный потенциал связан с диффузией только ионов калия.
Б. Мембранный потенциал связан с диффузией и ионов натрия, и ионов калия.
В. Мембранный потенциал связан с диффузией обоих ионов на фоне активного их транспорта Nа + /К + -насосом.
На рисунке представлены факторы, необходимые для установления нормального мембранного потенциала покоя -90 мВ.
Вклад калиевого диффузионного потенциала. Предположим, что единственным движением ионов через мембрану является диффузия ионов калия с помощью открытых каналов между символами калия (К+) внутри и снаружи мембраны. В связи с высоким отношением концентраций ионов калия внутри и снаружи (35 : 1) соответствующий этому отношению потенциал Нернста равен -94 мВ (результат умножения логарифма 35, равного 1,54, на -61 мВ). Следовательно, если бы единственным фактором, создающим потенциал покоя, являлись ионы калия, потенциал внутри волокна был бы равен -94 мВ, как показано на рисунке.
Вклад диффузии натрия через мембрану нерва. На рисунке ситуация дополнена небольшой проницаемостью мембраны нервного волокна для ионов натрия, связанной с незначительной диффузией ионов натрия через каналы К+/Nа+-утечки. Отношение концентраций ионов натрия внутри и снаружи мембраны составляет 0,1, следовательно, расчетный потенциал Нернста внутри мембраны равен +61 мВ. Но на рисунке показан также потенциал Нернста для диффузии ионов калия, равный -94 мВ. Как они взаимодействуют друг с другом, и каков будет суммарный потенциал? Для ответа нужно использовать уравнение Гольдмана, приведенное ранее.
Интуитивно можно предположить, что если мембрана высокопроницаема для ионов калия и лишь слегка проницаема для ионов натрия, то диффузия ионов калия вносит гораздо больший вклад в создание мембранного потенциала, чем диффузия ионов натрия. В норме проницаемость мембраны нервного волокна для ионов калия примерно в 100 раз выше, чем для ионов натрия. Использование этого значения в уравнении Гольдмана дает потенциал внутри мембраны около -86 мВ, что весьма близко к величине калиевого потенциала, показанного на рисунке.
Вклад натрий-калиевого насоса. Na+/K+-Hacoc обеспечивает дополнительный вклад в величину потенциала покоя за счет постоянного выкачивания 3 ионов натрия наружу в обмен на закачивание внутрь 2 ионов калия, что является причиной постоянной потери положительных зарядов с внутренней стороны мембраны. Это увеличивает степень электроотрицательности внутри мембраны примерно на -4 мВ в дополнение к создаваемой простой диффузией. Следовательно суммарный мембранный потенциал при совместном действии всех факторов составляет -90 мВ.
Итак, одни только диффузионные потенциалы, связанные с диффузией ионов калия и натрия, дают величину мембранного потенциала около -86 мВ, причем это значение почти полностью определяется диффузией ионов калия. Кроме того, постоянное функционирование электрогенного Na+/K+-Hacoca вносит дополнительные -4 мВ, давая суммарный мембранный потенциал, равный -90 мВ.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Мембранный потенциал и электрохимический градиент
• Мембранный потенциал возникает за счет электрохимического градиента, который существует по обеим сторонам мембраны, селективно проницаемой для ионов
• Величина мембранного потенциала как функции концентрации ионов рассчитывается по уравнению Нернста
• В клетке поддерживается отрицательное значение мембранного потенциала покоя. При этом внутренняя среда клетки, по сравнению с внешней, характеризуется несколько большим отрицательным зарядом
• Существование мембранного потенциала является необходимым условием генерации электрических сигналов, а также направленного транспорта ионов через мембрану
Важным свойством клеток является способность поддерживать такие внутриклеточные концентрации метаболитов, которые существенно отличаются от их содержания во внеклеточной среде. В случае ионов, различия в их концентрации по обеим сторонам мембраны приводят к различиям в электрическом заряде: внутриклеточная среда заряжена несколько более отрицательно, чем среда снаружи клетки. Совместное действие разности зарядов и концентраций проводит к возникновению электрохимического градиента. Электрохимический градиент поддерживается за счет действия селективных каналов и белков переносчиков в плазматической мембране.
Для того чтобы понять, каким образом возникает электрохимический градиент, вначале рассмотрим простой случай, когда мембрана оказывается проницаемой только для одного вида ионов. На рисунке ниже представлены два компартмента, А и В, разделенные тонкой мембраной. Эти компартменты содержат раствор КС1 разной концентрации. В растворе хлорид калия диссоциирован на гидратированные ионы К+ и Cl-. Поскольку оба компартмента содержат эквимолярные концентрации ионов, то каждый обладает нейтральным зарядом.
Если бы мембрана была непроницаема для ионов, то величина ее электрического потенциала, измеренная с помощью вольтметра, равнялась бы нулю.
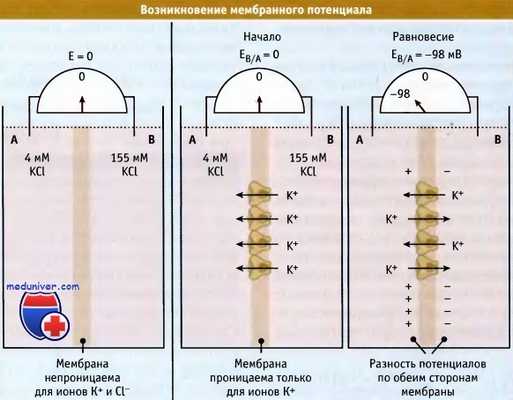
Селективное передвижение ионов через мембрану вызывает изменение мембранного потенциала.
Теперь рассмотрим случай, когда мембрана проницаема только для ионов калия (например, когда в мембране находятся К+-каналы). Диффузия растворенных веществ по градиенту концентрации является энергетически выгодным процессом (выражается в виде отрицательной величины разности энергии AG). Поэтому ионы К+ будут диффундировать в сторону более низкой их концентрации, т. е. из компартмента В в компартмент А. При этом распределение заряда на мембране будет меняться. По мере накопления в компартменте А положительно заряженных ионов, возрастают силы отталкивания между ними. Эти силы затрудняют переход ионов К+ в компартмент А.
Когда в системе достигается электрохимическое равновесие, градиенты концентрации и электрических зарядов взаимно уравновешиваются, и движение ионов К+ через мембрану прекращается. При этом транспорт ионов К+ из одного компартмента сдерживается их транспортом из другого компартмента.
Однако в компартменте А содержится больше положительно заряженных ионов, чем в компартменте В. Этот избыток ионов К+ (в компартменте А) взаимодействует с избытком ионов Cl- (в компартменте В) через тонкую мембрану, в результате чего по обеим ее сторонам выстраиваются электрические заряды. Разница зарядов по обеим сторонам мембраны выражается в виде разности потенциалов и называется мембранный потенциал. Равновесный (мембранный) потенциал компартмента В по отношению к компартменту А имеет отрицательное значение.
Этот пример иллюстрирует необходимость наличия двух условий, необходимых для возникновения мембранного потенциала клетки, не равного нулю:
• различные концентрации ионов по обеим сторонам мембраны, которые приводят к разделению зарядов и
• мембрана, обладающая селективной проницаемостью по крайней мере к одному виду ионов.
Поэтому величина мембранного потенциала является функцией концентрации ионов. В состоянии равновесия эту функцию для ионов X можно выразить количественно с помощью уравнения Нернста:
• Е — равновесный потенциал (в вольтах)
• R — универсальная газовая постоянная (2 кал моль -1 К -1 )
• Т — абсолютная температура (К; 37 °С = 307,5 К)
• z — валентность ионов (электрический заряд)
• F — число Фарадея (2,3 х 10 4 кал вольт -1 моль -1 )
• [Х]А — концентрация свободных ионов X в компартменте А
• [Х]в — концентрация свободных ионов X в компартменте В
В формировании мембранного потенциала в клетках животных, главным образом, участвуют ионы К+, Na+ и Cl-. Ионы Са2+ и Mg2+ в меньшей степени участвуют в формировании мембранного потенциала покоя. Плазматическая мембрана обладает селективной проницаемостью к перечисленным ионам (т. е. мембрана содержит ионные каналы, селективные к каждому типу ионов). Это обстоятельство, а также мембранная проницаемость (Р) для каждого иона учитывается в уравнении Гольдмана-Ходжкина-Каца, которое представляет собой расширенную форму уравнения Нернста.
Для основных ионов это уравнение выражает мембранный потенциал как функцию их проницаемости и концентрации внутри (i) и снаружи (о) клетки:
Величина отрицательного мембранного потенциала покоя зависит от типа клеток и колеблется от -200 мВ до -20 мВ. В клетках млекопитающих мембранный потенциал покоя в основном создается при работе К+-каналов и ионного насоса, который называется Na+/К+-АТФаза. Основной вклад в формирование отрицательного мембранного потенциала вносит небольшой поток ионов К+ через плазматическую мембрану. Этот поток осуществляется через К+-каналы, лишенные воротного механизма (т. н. калиевые каналы покоя).
В отличие от большинства других К+-каналов, которым необходим сигнал для открытия, эти каналы в клетке, обладающей определенным потенциалом покоя, открыты постоянно. В покоящейся клетке также открыты несколько каналов для других ионов. Движение ионов К+ из клетки, по направлению электрохимического градиента, помогает клеточному содержимому поддерживать отрицательный заряд. Пока мы не знаем всех источников ионов калия, которые участвуют в этом процессе. В некоторых клетках, например у растений и бактерий, а также в митохондриях, мембранный потенциал покоя создается за счет градиента протонов, а не ионов К+.
Для того чтобы происходила диффузия ионов К+ из клетки через К+-каналы, их концентрация в клетке должна быть выше, чем в окружающей среде. Градиент концентрации создается в результате работы Na+/К+-АТФа-зы, которая закачивает в клетку два иона калия на каждые три иона натрия, которые этот ионный насос удаляет из клетки. Поэтому насос функционирует как генератор заряда: удаляется больше электрических зарядов, чем привносится к клетку. Таким образом, наряду с K+-каналами, лишенными воротного механизма, Na+/К+-АТФазы участвуют в создании отрицательного внутриклеточного потенциала. Если происходит инактивация Na+/K+-АТФаз, то концентрации ионов Na+ и К+ по обе стороны мембраны уравниваются. Это происходит потому, что липидный бислой очень плохо пропускает ионы. Иными словами, без прохождения первичных процессов активного транспорта с участием Na+/К+-АТФаз значение мембранного потенциала равнялось бы нулю.
Мембранный потенциал покоящейся клетки представляет собой довольно постоянную величину. Однако при связывании лигандов, механическом стрессе или при изменении электрического заряда происходит открытие специфических ионных каналов, и мембранный потенциал изменяется. Если ионные каналы находятся под контролем электрического заряда, то изменения мембранного потенциала влияют на прохождение через них ионов. Открытие и закрытие канала контролируются воротным механизмом (гейтингом). Мембранный потенциал зависит от тех ионов, для которых каналы в основном, открыты. Например, при открытии Na+- или Са2+-каналов происходит деполяризация мембраны.
При этом соответствующие ионы начинают поступать в клетку в направлении их электрохимического градиента. Это приводит к тому, что мембранный потенциал становится более положительным. Напротив, при реполяризации мембраны (гиперполяризации) потенциал становится еще более отрицательным. Это происходит при открытии калиевых каналов и выходе из клетки ионов К+ в направлении градиента, что и влечет за собой увеличение отрицательного мембранного потенциала. Движение ионов по ионным каналам происходит быстро и исчисляется миллисекундами. Для изменения мембранного потенциала достаточны лишь незначительные различия в концентрации ионов по сторонам мембраны, и основная концентрация их в клетке не меняется.
Поток лишь 10 -12 моль К+ через 1 см2 мембраны приводит к ее быстрой гиперполяризации и к установлению мембранного потенциала, равного -100 мВ. Локальное передвижение относительно небольших зарядов через мембрану позволяет цитозолю и внеклеточной среде оставаться электрически нейтральными и сводит к минимуму электрическое отталкивание зарядов.
С энергетической точки зрения, мембранный потенциал представляет собой некий энергетический резервуар, энергию которого можно использовать для выполнения определенной работы. По расположению отрицательно заряженных ионов в цитозоле и положительно заряженных на наружной стороне мембраны, клетка напоминает электрический конденсатор или батарею, т. е. приспособление, способное сохранять электрическую энергию и служить ее источником. Энергия высвобождается в виде ионов, мигрирующих по направлению их электрохимического градиента, и может использоваться в процессах транспорта других ионов или метаболитов против градиента концентрации.
2_1 Мембранный потенциал покоя кратко (доцент Сазонов В.Ф.)
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое "животное электричество"? Откуда в организме берутся "биотоки"? Как живая клетка, находящаяся в водной среде, может превратиться в "электрическую батарейку"?
- На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов (заряженных частиц - ионов) создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
- На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка - нейрон.
В основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы - это понять, каким образом перемещаются электрические заряды в нервных клетках и как за счёт этого на мембране появляется электрический потенцил. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов - формирование потенциала покоя.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический потенциал на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя - это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
"Потенциал" - это возможность, он сродни понятию "потенция". Электрический потенциал мембраны - это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы - ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные - положительно (+).
Таблица 1. Физико-химические свойства ионов натрия и калия
Координационное число
Равновесный потенциал, мВ
Предельная температура гидратации, ºС
Однако, и Na + , и К + потенциально способны перемещаться под действием электрических сил туда, где преобладают не положительные, а отрицательные электрические заряды (отрицательно заряженные частицы - анионы).
Итак, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы с помощью электрических сил.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами - химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках - это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать "плюсики", т.е. положительно заряженные ионы, а внутри - "минусики", т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки - отрицательные "персонажи". ((
Сущность
Сущность потенциала покоя - это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки - "отрицательность", а снаружи - "положительность".
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны, (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях - и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий "хочет" войти в клетку, но "не может", т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне - 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку любые положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Попробую рассказать коротко, откуда берётся мембранный потенциал покоя в нервных клетках - нейронах. Ведь, как всем теперь известно, наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц - анионов и недостаток положительных частиц - катионов.
И вот тут исследователя и студента поджидает одна из логических ловушек: внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот - из-за потери некоторого количества положительных частиц (катионов).
И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов - катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомню, что это ионы натрия - Na + и калия - K + . Удивительно, но оба этих иона покидают клетку, хотя и разными путями.
С чего начать, чтобы не сбиться и не запутаться? Начнём с калия.
Чтобы легче было запомнить, образно можно сказать так:
"Клетка любит калий!"
или: "клетка копит калий!" (К + )
и "ненавидит натрий" (Na + ).
И что же она, клетка, делает с этими ионами? Она их обменивает друг на друга! С помощью специального обменного насоса.
Натрий-калиевый насос
Дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют "собственный" натрий клетки на наружный "чужой" калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Итак: "Клетка любит калий!" (Хотя об истинной любви здесь, разумеется, не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его итак уже полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия (любимого). Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов при формировании мышечного волокна потенциал их мембраны изменяется от -10 до -70 mV, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках (ММСК) костного мозга человека искусственная деполяризация ингибировала дифференцировку клеток (Fischer-Lougheed J., Liu J.H., Espinos E. et al. Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology 2001; 153: 677-85; Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology 1998; 510: 467-76; Sundelacruz S., Levin M., Kaplan D.L. Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. Plos One 2008; 3 ).
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка "заряжается любовью".
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Как ни странно, но результат этих двух видов любви - пустота!
Именно она, пустота, создаёт в клетке отрицательный электрический потенциал - потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал - возможность притягивать внутрь клетки любые положительно заряженные частицы (ионы).
2. Ионный натриевый химический потенциал - возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какого-нибудь другого вещества).
3. Ионный калиевый химический потенциал - возможновть выталкивать из клетки ионы калия (и именно калия, а не какого-нибудь другого вещества).
4. Дефицит натрия (Na + ) в клетке.
5. Избыток калия (K + ) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь "полезет" этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выровнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы "не замечают" друг друга, они реагируют только "на самих себя". Т.е. натрий реагирует на концентрацию натрия же, но "не обращает внимания" на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и "не замечает" натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно - концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий "хочет" снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко, и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
От химического - к электрическому
А теперь - самое главное, следите за излагаемой мыслью! Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и "плюсики" (положительные заряды). На их месте в клетке остаются "минусы" (отрицательные заряды). Это и есть мембранный потенциал покоя!
Мембранный потенциал покоя - это дефицит положительных зарядов (положительно заряженных ионов) внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Заключение
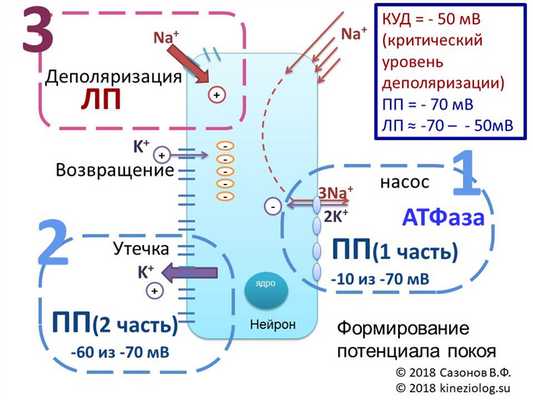
Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Потенциал покоя - отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть - это примерно -10 милливольт, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает "плюсиков" с натрием, чем закачивает обратно с калием).
2. Вторая часть - это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до -70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в -90 милливольт. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне -70 милливольт.
Обратите внимание на то, что для создания потенциала покоя нужны затраты энергии. Эти затраты производятся ионными насосами, которые обменивают "свой" внутренний натрий (ионы Na + ) на "чужой" внешний калий (K + ). Вспомним, что ионные насосы являются ферментами АТФазами и расщепляют АТФ, получая из неё энергию на указанный обмен ионов разного типа друг на друга.Тут очень важно понять, что с мембраной "работают" сразу 2 потенциала: химический (концентрационный градиент ионов) и электрический (разность электрических потенциалов по разные стороны мембраны). Ионы перемещаются в ту или иную сторону под действием обеих этих сил, на которые и тратится энергия. При этом один из двух потенциалов (химический или электрический) уменьшается, а другой увеличивается. Разумеется, если рассматривать электрический потенциал (разность потенциалов) отдельно, то не будут учитываться "химические" силы, перемещающие ионы. И тогда может сложиться неверное впечатление о том, что энергия на движение ионо берётся как бы ниоткуда. Но это не так. Необходимо рассматривать обе силы: химическую и электрическую. При этом крупные молекулы с отрицательными зарядами, находящиеся внутри клетки играют роль "статистов", т.к. их не перемещают через мембрану ни химические, ни электрические силы. Поэтому эти отрицательные частицы обычно и не рассматривают, хотя они существуют и именно они обеспечивают отрицательную сторону разности потенциалов между внутренней и наружной сторонами мембраны. А вот шустрые ионы калия, как раз способны к перемещению, и именно их утечка из клетки под действием химических сил создаёт львиную долю электрического потенциала (разности потенциалов). Ведь именно ионы калия перемещают на наружную сторону мембраны положительные электрические заряды, будучи положительно заряженными частицами.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки "лишнего" калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть "мембранный потенциал покоя". Он измеряется внутри клетки и составляет обычно -70 мВ.
Выводы
Говоря образно, "мембрана превращает клетку в "электрическую батарейку" с помощью управления ионными потоками".
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше "плюсиков" (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки - неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это - разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Видео: Мембранный потенциал покоя (Resting membrane potential)
Читайте также: