Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
Добавил пользователь Владимир З. Обновлено: 29.01.2026
Герпесвирусы (от греч. herpes- ползучий) -семейство крупных оболочечных ДНК-содержащих вирусов, вызывающих разнообразные инфекции.
Семейство Herpesviridae включает три подсемейства, отличающиеся по структуре генома, тканевому тропизму, цитопатологии и локализации латентной инфекции:
- подсемейство Alphaherpesvirinae - вирусы герпеса (ВПГ-1, ВПГ-2, VZV): для этой группы характерен быстрый рост. Вирусы размножаются в эпителиальных клетках, вызывая цитолитическое действие. В нейронах вызывают латентную, персистирующую инфекцию;
- подсемейство Betaherpesvirinae - вирусы герпеса (ЦМВ, ГВЧ-6, ГВЧ-7): для этой группы характерен медленный рост (латентная инфекция) в клетках эпителия слюнных желез, в гландах, почках, лимфоцитах. Вирусы оказывают цитомегалическое действие (ЦМВ) и лимфопролиферативное действие;
- подсемейство Gammaherpesvirinae - вирусы (ВЭБ) растут в лимфобластоидных клетках, оказывают лимфопролиферативное действие. Вызывают латентную инфекцию в лимфоидной ткани, лимфоцитах, эпителиальных клетках рта и глотки, слюнных желез. ВЭБ вызывает размножение В-лимфоцитов и персистирует в них.
Структура. Вирион герпесвируса имеет овальную форму диаметром 150-200 нм.
В центральной части вириона находится двунитевая линейная ДНК, окруженная икосаэдрическим капсидом, состоящим из 162 капсомеров. Снаружи вирус окружает оболочка с гликопротеиновыми шипами, сформированными из внутреннего слоя ядерной мембраны клетки. Пространство между капсидом и оболочкой называется тегумент (содержит вирусные белки и ферменты, необходимые для инициации репликации). Геном - двунитевая линейная ДНК. Она состоит: у ВПГ и ЦМВ - из двух фрагментов: короткого S и длинного L, каждый из которых у ВПГ заключен между двумя наборами инвертированных повторов, позволяющим геному рекомбинировать с образованием 4 изомеров; у VZV ДНК также состоит из двух фрагментов: короткого S и длинного L, но содержит один набор инвертированных повторов, поэтому формируется две изомерные формы.
Репродукция.
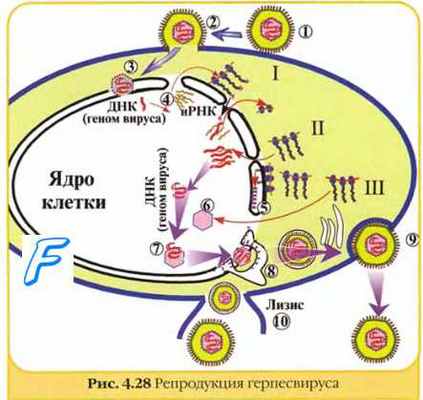
После прикрепления к рецепторам клетки оболочка вириона сливается с клеточной мембраной (1, 2). Освободившийся нуклеокапсид (3) доставляет в ядро клетки ДНК вируса. Далее происходит транскрипция части вирусного генома (с помощью клеточной ДНК-зависимой РНК-полимеразы); образовавшиеся иРНК (4) проникают в цитоплазму где происходит синтез (трансляция) самых ранних альфа-белков (I), обладающих регулирующей активностью. Затем синтезируются ранние бета-белки (П) - ферменты, включая ДНК-зависимую ДНК-полимеразу и тимидинкиназу, участвующие в репликации геномной ДНК вируса. Поздние гамма- белки (Ш) являются структурными белками, включая капсид и гликопротеины (А, В, С, D, Е, F, G, X). Гликопротеины диффузно прилегают к ядерной оболочке (5). Формирующийся капсид (6) заполняется вирусной ДНК и почкуется через модифицированные мембраны ядерной оболочки (8). Перемещаясь через аппарат Гольджи, вирионы транспортируются через цитоплазму и выходят из клетки путем экзоцитоза (9) или лизиса клетки (10).
Вирус простого герпеса относится к семейству Herpesviridae, роду Simplexvirus. Вызывает простой герпес (herpes simplex), характеризующийся везикулезными высыпаниями на коже, слизистых оболочках, поражением центральной нервной системы и внутренних органов, а также пожизненным носительством (персистенцией) и рецидивами болезни. Вирус простого герпеса включает два типа: ВПГ-1 и ВПГ-2; распространен повсеместно, поражает большую часть населения Земли и существует в организме в латентной форме до момента реактивации.
ВПГ-1 поражает преимущественно область рта, глаз, ЦНС, а ВПГ-2 - гениталии, за что и получил название генитального штамма .
Оглавление темы "Вирусология. Репродукция вирусов. Генетика вирусов.":
1. Вирусология. История вирусологии. Шамберлан. Ру. Пастер. Ивановский.
2. Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
3. Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
4. Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
5. Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
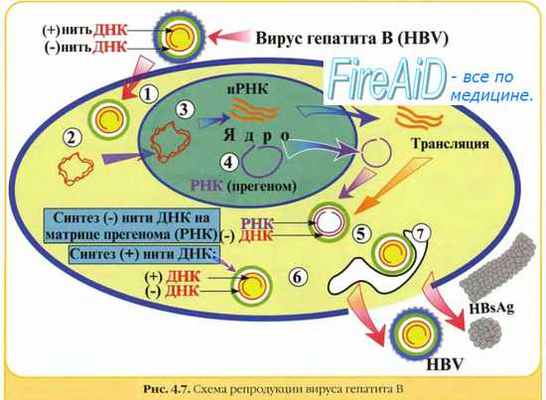
7. Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
8. Генетика вирусов. Характеристика вирусных популяций. Генофонд вирусных популяций.
9. Мутации вирусов. Спонтанные мутации вирусов. Индуцированные мутации вирусов. Проявление мутаций вирусов в фенотипе.
10. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами. Обмен фрагментами генома вирусами. Антигенный шифт.
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко.
Ранняя стадия репродукции герпесвирусов . В ранней стадии синтезируются «ранние белки», кодируемые проксимальной третью молекулы ДНК. Они проявляют регуляторные свойства, включая активацию транскрипции других участков вирусного генома, кодирующих ДНК-полимеразу и ДНК-связывающие белки.
Поздняя стадия репродукции герпесвирусов . В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов).
Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов - формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
Поксвирусы. Репродукция поксвирусов.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью осуществляется вирусными полимеразами.
Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз, так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНК-полимеразу, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий.
В репродуктивном цикле выделяют три стадии- начальную, раннюю и позднюю.
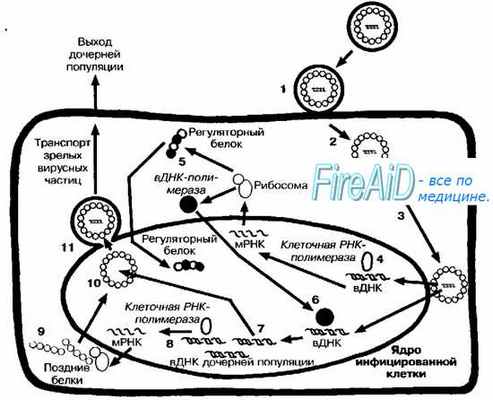
Рис. 5-4. Репликативный цикл ДНК-содержащих вирусов (на примере репродукции вируса герпеса). После адсорбции (1) вирус проникает в клетку путём слияния с мембраной (2). Нуклеокапсид транспортируется к ядерной оболочке (3), и вирусная ДНК (вДНК) проникает в ядро клетки, где начинается её транскрипция клеточной ДНК-зависимой РНК-полимеразой (4). Вначале транскрибируются «ранние гены». В результате трансляции «ранней порции» вирусного генома синтезируются «ранние белки» (5), включая регуляторные, матричные и вирусные полимеразы. Вирусная полимераза проникает в геном клетки (6), где запускает синтез молекул ДНК дочерних популяций (7). Часть вирусной ДНК («поздние гены») дочерних популяций транскрибируется клеточной РНК-полимеразой (8), что приводит к синтезу «поздних белков» (9), необходимых для сборки дочерних популяций (10). Последние покидают ядро, отпочковываясь от его мембраны (11), фрагменты которой содержат в составе своей обопочки.
Начальная стадия репродукции поксвирусов запускается сразу же после раздевания вируса и выхода вирусной ДНК в цитоплазму.
Ранняя стадия репродукции поксвирусов . На этом этапе транскрибируется примерно половина вирусной ДНК. Синтезируются ферменты, кодируемые «ранними генами» и участвующие в репликации вирусной ДНК. Параллельно в небольшом количестве образуются структурные белки.
Поздняя стадия репродукции поксвирусов совпадает с началом репликации ДНК (что переключает механизмы транскрипции на считывание второй половины генома). Регуляторные белки блокируют трансляцию «ранней мРНК» и запускают синтез поздних (структурных) белков. Сборка вирионов осуществляется только в цитоплазме посредством реакций мембранного синтеза. Высвобождение зрелых популяций сопровождается лизисом клетки.
НЕКОТОРЫЕ СВЕДЕНИЯ О МОРФОЛОГИИ ГЕРПЕС-ВИРУСОВ И ИХ СВОЙСТВАХ
Дальневосточный научный центр физиологии и патологии дыхания Сибирского отделения РАМН,
В настоящей работе представлены литературные сведения, характеризующие строение вирусов простого герпеса и их механизм взаимодействия с клетками-мишенями.
SUMMARY M.T.Lutsenko, I.N.Gorikov
SOME DATA ABOUT HERPES-VIRUSES
MORPHOLOGY AND THEIR PROPERTIES
The reference data characterizing the structure of simple herpes viruses and the mechanism of their interaction with cells-targets are given in the work.
Key words: virus, herpes.
В вирионе герпеса выявляют 3 компонента: 1) нук-леонд, локализующийся в центральной части; 2) кап-сид, покрывающий нуклеоид и представленный капсомерами; 3) оболочки, которые окружают эти структурные образования . Оболочка вирионов герпеса обычно сохраняет гексагональную форму. Диаметр оболочки составляет от 170 до 210 нм. Встречаются два и более нуклеокапсида, имеющих общую оболочку. Часто обнаруживаются вирусные частицы, которые не имеют оболочки. Капсид обычно гексагональной формы. Каждая грань капсида является равносторонним треугольником, состоящим из 15 субъединиц (интервал между субъединицами 3 нм). При использовании метода негативного контрастирования установлено, что капсид герпесвирусов является икосаэдром. Капсомеры - это полые структуры, имеющие на поперечном срезе пента- и гексагональное строение. Ребро икосаэдра представлено 5 капсомерами. 12 вершин образованы одним из капсомеров и окружены пятью соседними. Другие капсомеры граней треугольников также ограничены пятью соседними. Капсомер сохраняет форму удлиненной призмы. Его размеры составляют 9,5 х 12,5 нм. На поперечном срезе вершины икосаэдра они имеют пентагональную форму. Остальные капсомеры поверхности капсида имеют гексогональную форму с внутренним отверстием до 4 нм. Таким образом, капсид вириона герпеса представлен 162 капсомерами, которые упакованы в симметричном порядке, в соотношении 5:3:2 (рис. 1). При проведении электронной микроскопии преобладают вирионы (с оболочкой или без нее), в центральную часть которых не проникает фосфорно-вольфрамовая кислота. Эти вирионы
условно названы «полными», то есть они содержат нуклеоид. Одновременно идентифицируются вирионы, у которых фосфорно-вольфрамовая кислота определяется в их центральной части. Данный морфологический факт позволяет назвать их «пустыми» ви-рионами и предположить отсутствие у них нуклеоида. У таких вирионов обычно четко контурируется капсид. В его составе выявляется до 24 капсомеров. По мнению автора , ограниченное оболочкой капсида гексагональное пространство, в котором четко контурируется фосфорно-вольфрамовая кислота, имеет средний размер 78 нм (рис. 2).
Рис. 2. Вирус простого герпеса. Срез зараженной клетки фибробласта. Незрелые вирионы в ядре клетки (по А.Ф.Бочарову). Увеличение >
Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко.
Ранняя стадия репродукции герпесвирусов. В ранней стадии синтезируются «ранние белки», кодируемые проксимальной третью молекулы ДНК. Они проявляют регуляторные свойства, включая активацию транскрипции других участков вирусного генома, кодирующих ДНК-полимеразу и ДНК-связывающие белки.
Поздняя стадия репродукции герпесвирусов. В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов).
Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов — формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
Поксвирусы. Репродукция поксвирусов.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
• Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью осуществляется вирусными полимеразами.
• Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз, так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНК-полимеразу, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий.
В репродуктивном цикле выделяют три стадии— начальную, раннюю и позднюю.
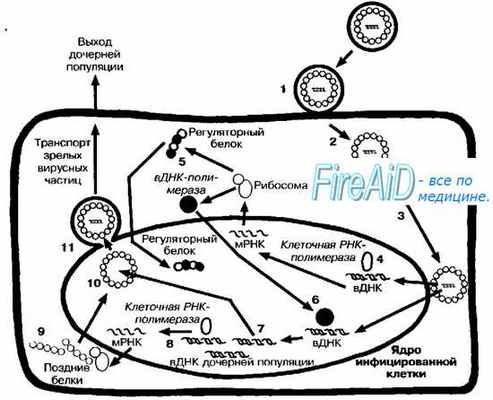
Рис. 5-4. Репликативный цикл ДНК-содержащих вирусов (на примере репродукции вируса герпеса). После адсорбции (1) вирус проникает в клетку путём слияния с мембраной (2). Нуклеокапсид транспортируется к ядерной оболочке (3), и вирусная ДНК (вДНК) проникает в ядро клетки, где начинается её транскрипция клеточной ДНК-зависимой РНК-полимеразой (4). Вначале транскрибируются «ранние гены». В результате трансляции «ранней порции» вирусного генома синтезируются «ранние белки» (5), включая регуляторные, матричные и вирусные полимеразы. Вирусная полимераза проникает в геном клетки (6), где запускает синтез молекул ДНК дочерних популяций (7). Часть вирусной ДНК («поздние гены») дочерних популяций транскрибируется клеточной РНК-полимеразой (8), что приводит к синтезу «поздних белков» (9), необходимых для сборки дочерних популяций (10). Последние покидают ядро, отпочковываясь от его мембраны (11), фрагменты которой содержат в составе своей обопочки.
Начальная стадия репродукции поксвирусов запускается сразу же после раздевания вируса и выхода вирусной ДНК в цитоплазму.
Ранняя стадия репродукции поксвирусов. На этом этапе транскрибируется примерно половина вирусной ДНК. Синтезируются ферменты, кодируемые «ранними генами» и участвующие в репликации вирусной ДНК. Параллельно в небольшом количестве образуются структурные белки.
Поздняя стадия репродукции поксвирусов совпадает с началом репликации ДНК (что переключает механизмы транскрипции на считывание второй половины генома). Регуляторные белки блокируют трансляцию «ранней мРНК» и запускают синтез поздних (структурных) белков. Сборка вирионов осуществляется только в цитоплазме посредством реакций мембранного синтеза. Высвобождение зрелых популяций сопровождается лизисом клетки.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аде
Оглавление темы "Вирусология. Репродукция вирусов. Генетика вирусов.":
1. Вирусология. История вирусологии. Шамберлан. Ру. Пастер. Ивановский.
2. Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
3. Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
4. Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
5. Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
6. Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
7. Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
8. Генетика вирусов. Характеристика вирусных популяций. Генофонд вирусных популяций.
9. Мутации вирусов. Спонтанные мутации вирусов. Индуцированные мутации вирусов. Проявление мутаций вирусов в фенотипе.
10. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами. Обмен фрагментами генома вирусами. Антигенный шифт.
Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Ранняя стадия репродукции. Вирусная ДНК проникает в ядро клетки, где транскрибируется клеточной ДНК-зависимой РНК-полимеразой. При этом считываетеся, а затем транслируется часть вирусного генома («paнние гены»). В результате синтезируются «ранние белки» (регуляторные и матричные белки вирусные полимеразы).
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
Матричные белки необходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Поздняя стадия репродукции. На этом этапе происходит синтез нуклеиновых кислот вируса. Не вся вновь синтезированная вирусная ДНК упаковывается в вирионы дочерней популяции. Часть ДНК («поздние гены») используется для синтеза «поздних белков», необходимых для сборки вирионов. Их образование катализируют вирусные и модифицированные клеточные полимеразы.
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
Ранняя фаза репродукции. Вирусная ДНК («ранние гены») транскрибируется в ядре клетки. На одной из нитей ДНК реализуется транскрипция вирусной «ранней» мРНК. Механизмы транскрипции вирусной ДНК аналогичны считыванию информации с клеточной ДНК. Специфическая мРНК транслируется, начинается синтез ферментов, необходимых для образования дочерних копий ДНК. Синтез клеточной ДНК может временно усиливаться, но затем обязательно подавляется регуляторными белками вируса.
Поздняя фаза репродукции. В течение поздней фазы дочерняя вирусная ДНК продолжает активно транскрибироваться клеточными РНК-полимеразами, в результате чего появляются продукты поздних вирусспецифических синтезов. «Поздняя» мРНК мигрирует в цитоплазму и транслируется на рибосомах. В результате синтезируются капсидные белки дочерней популяции, которые транспортируются в ядро и собираются вокруг молекул дочерней ДНК новых вирусных частиц. Выход полных дочерних популяций сопровождается гибелью клетки.
- Читать далее "Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов."
Вирусы с дц ДНК
• Репликация генома у большинства ДНК-содержащих вирусов
происходит в клеточном ядре.
• Большинство ДНК-содержащих вирусов полностью полагаются на
синтетический аппарат клетки-хозяина. Однако вирусы с
крупными геномами (например, поксвирусы) могут сами
кодировать большую часть необходимых для этого белков.
3. Класс I:
• Вирусы, содержащие двуцепочечную ДНК
для репликации попадают в ядро клетки, так как им требуется
клеточная ДНК-полимераза. Также репликация ДНК этих вирусов
сильно зависит от стадии клеточного цикла. В некоторых случаях
вирус может вызывать деления клетки, что может приводить к
раковому перерождению. Примерами таких вирусов
являются Herpesvirales, Adenoviridae, Papillomaviridae и Polyomaviri
dae.
• У представителей семейства Poxviridae геномная ДНК
реплицируется не в ядре.
4. Herpesvirales
Herpesvirales (лат., от греч. herpes — ползучий) — порядок вирусов
Крупный линейный геном
дцДНК
икосаэдрический капсид(T=16)
белковый тегумент и липидная оболочка, в которой расположены
мембранные белки
• Иммунносупресия организма хозяина
• Постоянно находятся в организме
• Онкогенны
5. Геном
• Представлет собой линейную двухцепочечную молекулу ДНК
• У герпесвируса человека 1-го типа она имеет размер 152 261 пара
нуклеотидов и содержит 77 генов, все из которых кодируют
белки; доля пар ГЦ составляет 68 %
• Геном герпесвируса человека 2-го типа состоит из 154 746 пар
нуклеотидов и несёт 77 генов; доля пар ГЦ составляет 70 %
6. Инфицирование
Вид герпесвируса человека
Герпесвирус человека тип 1
Подсемейств
Вызываемая болезнь
о вирусов
Вирус простого герпеса первого типа (ВПГ- Альфагерпес Оральный и генитальный герпес, но чаще оральный
1, Herpes simplex virus-1, HSV-1)
вирусы
(герпетический стоматит, губной герпес)
Название вируса
Герпесвирус человека тип 2
Вирус простого герпеса второго типа (ВПГ- Альфагерпес Оральный и генитальный герпес, но чаще генитальный и
2, Herpes simplex virus-2, HSV-2)
вирусы
вагинальный герпес
Герпесвирус человека тип 3
Вирус ветряной оспы (Varicella-zoster virus, Альфагерпес
Ветряная оспа , опоясывающий лишай (зостер, zoster)
VZV)
вирусы
Герпесвирус человека тип 4
Инфекционный мононуклеоз, лимфома Беркитта, лимфома
Вирус Эпштейна — Барр (ВЭБ, Epstein-Barr Гаммагерпес ЦНС у больных с иммунодефицитным синдромом,
virus, EBV)
вирусы
посттрансплантантный лимфопролиферативный синдром
назофарингеальная карцинома
Герпесвирус человека тип 5
Инфекционный мононуклеоз, ретинит, гепатит, увеличение
Бетагерпесви
органов брюшной полости, воспаление слюнных желез (так
русы
называемое слюнотечение)
Цитомегаловирус человека
•Розеоловирусы: Human herpesvirus 6A,
•Герпесвирус человека тип 6 HHV-6A
•Human herpesvirus 6B, HHV-6B
Шестая болезнь — детская розеола (розеола инфантум,
Бетагерпесви
roseola infantum) или экзантема (экзантема субитум,
русы
exanthem subitum)
Герпесвирус человека тип 7
Розеоловирус (Human herpesvirus 7, HHV-7)
Бетагерпесви Вероятная причина синдрома хронической усталости (СХУ)[3].
русы
Часто сосуществует с вирусом герпеса 6 типа
Герпесвирус человека тип 8
Герпесвирус, ассоциированный с саркомой
Гаммагерпес Саркома Капоши, первичная лимфома серозных полостей,
Капоши(Kaposi’s sarcoma-associated
вирусы
некоторые разновидности болезни Кастельмана
herpesvirus, KSHV)
8. Adenoviridae
Adenoviridae
• Аде́новирусы (лат. Adenoviridae) — семейство ДНК-содержащих
вирусов позвоночных, лишённых липопротеиновой оболочки.
• диаметр 70—90 нм, содержат единичную двухцепочечную
молекулу ДНК длиной 34-36 т.п.н., молекулярной массой 20—
29·106 Да.
• Наиболее известны аденовирусы, вызывающие острые
респираторные заболевания.
• На материале аденовирусов впервые было открыто явление
альтернативного сплайсинга
9. Вред и польза
• Большая часть серотипов аденовирусов приводят к инфекции
верхних дыхательных путей. Также зачастую аденовирусы бывают
причиной конъюнктивита, тонзиллита, отита. Также аденовирусы
40 и 41 серотипа могут вызывать гастроэнтерит.
• Не существует специфической антивирусной терапии для лечения
аденовирусной инфекции, и лечение в основном направлено на
устранение симптомов.
• Аденовирусы используются в качестве вирусного вектора для
генной терапии благодаря их способности размножаться в
реплицирующихся и нереплицирующихся клетках
• В Китае аденовирусы используются в лечении онкологических
заболеваний.
11. Papillomaviridae
Papillomaviridae
• Размеры вириона около 55 нм. Размножается в ядре
клеток-хозяев
• Причина возникновения доброкачесвенных опухолей
• Папилломавирусы имеют видоспецифические и некоторые
типоспецифические антигены.
• Не культивируются в клеточных системах.
• Вирус папилломы человека выявляется в кератоцитах в виде
эписомы, 50—200 копий на поражённую клетку.
12. Патогенез
• известно более 600 штаммов
папилломавирусов, обнаруженных у человека
• более 40 из них могут вызвать поражение
аногенитального тракта мужчин и женщин и появление
остроконечных кондилом.
• безвредны /вызывают бородавки /рак
• Единственный способ заболевания раком шейки матки
• Инфицируют базальный слой эпителия, причём наиболее поражённым участком является
зона перехода многослойного плоского эпителия в цилиндрический эпителий.
• В заражённой клетке вирус существует в двух формах:
Эписомальной
Интросомальной
13. Симптоматика
• Понижение иммунитета
• Длительный инкубационный период(от половины месяца до нескольких лет)
• Метод ПЦР имеет большую диагностическую значимость и позволяет идентифицировать отдельные типы
ВПЧ
• Скрининговый используется (Digene-тест) для быстрого выявления клинически значимых концентраций
высоконкогенных типов папилломавируса. Имеет высокую специфичность и точность диагностики.
Производится в комплексе с цитологическим исследованием шейки матки.
• Так как аногенитальные бородавки в 90 % случаев ассоциируются с другими урогенитальными инфекциями,
целесообразно проводить исследования на их наличие методом ПЦР
• Согласно исследованиям Центров по контролю и профилактике заболеваний США, естественный иммунитет
самостоятельно справляется с вирусом в организме в течение 2 лет в 90 % случаев[.
• Средствами удаления поражённых тканей могут быть:
хирургическое удаление;
электрокоагуляция
лазерокоагуляция углекислым или неодимовым лазером
криодеструкция жидким азотом
• Побочным эффектом может быть распространение вируса на прилегающие ткани, повреждённые в
процессе удаления поражённых участков.
14. Polyomaviridae
• вирионы небольшие,
диаметр порядка 40—50 нм, икосаэдрической формы
• не покрыты липидной оболочкой.
• Вирусы обычно онкогенные, часто находятся в организме хозяина
в латентном состоянии и не вызывают болезнь, но образуют
опухоли в случае иммунного дефицита хозяина.
• Корень «полиома» в названии вируса говорит о том, что вирусы
способны вызывать множественные опухоли.
• Инфекцию, вызываемую Human polyomavirus 1—4, сложно отличить от
инфекции, вызываемой Macaca mulatta polyomavirus 1
• Human polyomavirus 1 приводит к мягким респираторным инфекциям,
и поражает почки у пациентов со сниженной иммунной системой,
например, после трансплантации органов. Human polyomavirus 2
поражает клетки дыхательной системы, почек или мозга. Оба эти
вируса широко распространены в популяции, порядка 80 % взрослых
жителей США имеют антитела к этим вирусам.
• Human polyomavirus 5 описан в 2008 году, вызывает рак кожи Меркеля
• Human polyomavirus 8 описан в 2010 году, вызывает триходисплазию.
16. Поксвирусы
• Поксви́ русы или вирусы оспы (лат. Poxviridae, от англ. рох — оспа +
вирусы) — семейство вирусов животных, объединяющее самые
крупные ДНК-содержащие вирусы, вирионы которых имеют
кирпичеобразную или овоидную форму; включает виды, патогенные
для человека, например, возбудителей оспы, пустулезного дерматита.
• Геном, представленный двунитчатой линейной гантелеобразной
формы ДНК
• Двухслойный капсид, между слоями которого находятся боковые тела.
• Поверх нуклеокапсида расположена двухслойная липопротеидная
оболочка с воронкообразными фибрами.
17. Репродукция вируса
• Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется
более 100 различных белков, входящих в состав вирионов (большинство образует наружную
оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
• Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью
осуществляется белками, имеющимися в вирионе.
• Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз,
так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНКполимеразу, которая обеспечивает транскрипцию генов репликации в течение ранней и средней
стадий.
• В репродуктивном цикле выделяют три стадии:
• Ранняя стадия репродукции поксвирусов запускается сразу же после раздевания вируса и
выхода вирусной ДНК в цитоплазму. На этой стадии за счёт РНК-полимеразы вириона
транскрибируются гены, необходимые для репликации генома.
• На средней стадии транскрибируется примерно половина вирусной ДНК. Транслируются мРНК,
синтезированные в раннюю стадию. Транскрибируются средние гены.
• Поздняя стадия репродукции поксвирусов совпадает с началом репликации ДНК. Регуляторные
белки блокируют трансляцию ранней мРНК и запускают синтез поздних (структурных) белков.
Сборка вирионов осуществляется только в цитоплазме посредством реакций мембранного
синтеза. Упаковка генома в вирион идёт с затратой АТФ. Высвобождение зрелых популяций
сопровождается лизисом клетки.
18. Мимивирус
• Единственный опознанный вид Acanthamoeba polyphaga
mimivirus (APMV).
• До октября 2011 года, когда был описан ещё более крупный вирус
Megavirus chilensis, считалось, что мимивирус обладает самым
крупным капсидом среди всех известных вирусов — около 500 нм
в диаметре.
• В отличие от большинства других вирусов, мимивирус не проходит
через фильтр с диаметром пор 0,22 мкм
• Виден в световой микроскоп, по размерам он близок к небольшим
бактериям, таким как микоплазма. Кроме того, по сравнению с
другими вирусами и даже некоторыми бактериями, мимивирус
обладает более объёмным (около 1,2 миллионов пар
нуклеотидов) и сложноструктурированным геномом
• В свете нехватки точных данных о природе данного вируса его
открытие вызвало большой интерес в научных кругах. Один из
первооткрывателей мимивируса высказал предположение, что он
представляет собой недостающее звено между вирусами и
клеточными организмами. Имеется и более радикальное мнение,
согласно которому мимивирус представляет собой принципиально
новую форму жизни, не относящуюся к вирусам или бактериям.
19. Открытие и этимология
• Название «мимивирус» было дано этому вирусу как сокращение от
«мимикрирующий под микроб» (англ. mimicking microbe virus). Это связано с тем,
что некоторое время этот вирус считали микроорганизмом, а не вирусом, на
основании таких признаков, как крупный размер, наличие белковых нитей, похожих
на жгутики, и способность окрашиваться по методу Грама.
• APMV был впервые обнаружен в 1992 году в амёбе Acanthamoeba polyphaga, в
честь которой он и был назван, в ходе поисков возбудителей легионеллёза. Вирус
был обнаружен в препарате, окрашенном по методу Грама, и вследствие этого
ошибочно принят за грамположительную бактерию. Организм был назван
Bradfordcoccus в честь района, в котором была найдена амёба (Брадфорд, Англия).
После неудачных попыток культивирования и ПЦР-типирования этого организма с
помощью универсальных праймеров, узнающих гены бактериальной 16S рРНК,
образец пролежал в холодильнике 10 лет.
• Позднее он был передан во Францию, где были проведены дополнительные
исследования, позволившие заключить, что Bradfordcoccus на самом деле является
гигантским вирусом. Результаты работы были опубликованы в 2003 году в журнале
«Science»
20. Немного о геноме
• Ряд белков мимивируса, принимающих участие в репликации
генома, оказались гомологичными белкам других крупных
ядерно-цитоплазматических ДНК-содержащих вирусов, что
говорит об их общем происхождении.
• Большое количество мимивирусных белков не обнаруживают
сходства ни с одним известным в настоящее время белком.
• Геном кодирует значительное количество белков, напоминающих
эукариотические и бактериальные. По-видимому, эти гены были
приобретены мимивирусом вторично и происходят из геномов
хозяев вируса и их паразитов.
• обладает бо́ льшим объёмом генетической информации, чем как
минимум 30 организмов, имеющих клеточное строение
• Несмотря на большой размер генома, он используется довольно
эффективно: на долю кодирующей ДНК приходится около 90,5 % всего
объёма геном
• Открытые рамки считывания разделены между собой промежутками
примерно в 157 пар нуклеотидов.
• Две цепочки ДНК, названные R (англ. right — правая) и L (англ. left —
левая), кодируют примерно одинаковое количество генов (450 и 465
соответственно)
• Не онаружено признаков деградации, характерных для геномов
паразитических бактерий, таких как псевдогены и мобильные
генетические элементы. Содержание адениновых и тимидиновых
нуклеотидов довольно высоко — 72 %, что ведёт к повышенному
содержанию в белках аминокислот, кодируемых АТ-богатыми
кодонами (например, изолейцина, аспарагина и тирозина).
• Вблизи концов молекулы ДНК были обнаружены инвертированные
повторы длиной 617 пар нуклеотидов. Предполагают, что
комплементарное взаимодействие этих участков может приводить к
формированию Q-структуры — кольцевой ДНК с двумя небольшими
хвостами
• Есть признаки, соответствующие принципам ламаркизма. В условиях
пониженной конкуренции происходит репрессия некоторых генов. Это
изменение наследуется следующими поколениями вируса, в некоторых
случаях усиливаясь вплоть до полной инактивации этих генов.
Предположительно, эффект является следствием худшей репарации
менее используемых генов
• Мимивирус является одним из немногих дцДНК-вирусов, в геноме
которых обнаружена последовательность, кодирующая интеин. Интеин
представляет собой белковый домен, который катализирует своё
собственное вырезание из несущей молекулы и сшивание
образовавшихся концов. Такая последовательность присутствует в гене
ДНК-полимеразы B мимивируса
• Октамерная последовательность AAAATTGA была обнаружена
примерно перед половиной генов мимивируса в положении от −80 до
−50. Эта последовательность является TATA-подобным промоторным
элементом и узнаётся транскрипционным аппаратом вируса на ранней
стадии жизненного цикла. Другая вырожденная АТ-богатая
последовательность является поздним промотором
• 24-часовой литический (сопровождающийся лизисом клетки-хозяина) жизненный цикл
• Вирионы мимивируса, напоминающие бактерий своими размерами и присутствием
характерных полисахаридов на поверхности, поглощаются амёбой как пища в процессе
эндоцитоза
• Белковые нити частично лизируются в эндосомах, в результате чего капсид может
взаимодействовать с эндосомальной мембраной;
• Через 2 часа после инфицирования капсид раскрывается в районе звёздчатой структуры, и его
содержимое высвобождается в цитоплазму в результате слияния внутренней мембраны и
мембраны эндосомы
• После выхода коровой частицы (внутренней части нуклеокапсида) в цитоплазму, благодаря
наличию в ней вирусного аппарата транскрипции, начинается синтез вирусных мРНК. Эти мРНК
накапливаются внутри коровой частицы в виде гранул. Предполагают, что первыми под
действием вирусной РНК-полимеразы транскрибируются гены, которые находятся под
контролем AAAATTGA-промотора
• Через 4—5 часов после заражения вирусная ДНК выходит из коровой частицы и
деконденсируется, начинается репликация. В результате рядом с пустой оболочкой коровой
частицы формируется так называемая «вирусная фабрика» — место синтеза компонентов и
сборки вирусных частиц. Если в клетку проникли несколько вирусных частиц, то формируемые
ими «фабрики» сливаются в одну по мере разрастания;
• Через 6—9 часов после заражения можно наблюдать процессы сборки капсидов и упаковки в
них ДНК, которые происходят на периферии «вирусных фабрик». Необычным свойством
мимивируса является то, что ДНК упаковывается и выходит из капсида через два разных
отверстия
• Через 14—24 часа после инфекции происходит лизис клетки амёбы и высвобождение
вирионов, к этому моменту их накапливается в клетке более 300 единиц.
25. Патогенез
• Существует гипотеза, что мимивирус может вызывать у людей
некоторые формы пневмонии
• Мимивирус способен заражать человеческие макрофаги, проникая в
клетки в процессе фагоцитоза, и реплицироваться в них.
• В нескольких исследованиях у небольшого количества пациентов,
страдающих пневмонией, были обнаружены антитела против
мимивируса. Также был описан единичный случай развития
пневмонии у лаборанта, который работал с культурами этого вируса.
Содержание антител против мимивируса в его крови тоже было
повышенным
• Присутствие антител против вируса само по себе не говорит о его
патогенности, не исключено, что мимивирус просто обладает
сильными иммуногенными свойствами.
26. Дополнительная информация
• Есть вирус- сателлит
• Возможно он вирофаг
• Ближайшие родственники возможно
• микоплазмы и риккетсии
Вирус герпеса и репродукция
Оглавление темы “Вирусология. Репродукция вирусов. Генетика вирусов.”:
1. Вирусология. История вирусологии. Шамберлан. Ру. Пастер. Ивановский.
2. Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
3. Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
4. Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
5. Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
6. Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
7. Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
8. Генетика вирусов. Характеристика вирусных популяций. Генофонд вирусных популяций.
9. Мутации вирусов. Спонтанные мутации вирусов. Индуцированные мутации вирусов. Проявление мутаций вирусов в фенотипе.
10. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами. Обмен фрагментами генома вирусами. Антигенный шифт.
Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
Основные отличия репродуктивного цикла герпесвирусов от остальных ДНК-вирусов связаны с более сложной структурой генома. Адсорбция вирусов на клетках осуществляется через специфические рецепторы. После взаимодействия с рецепторами вирусная оболочка сливается с клеточной мембраной, а нуклеокапсид высвобождается в цитоплазму. Раздевание (депротеинизация) вирусного генома происходит на ядерной мембране, и вирусная ДНК оказывается в ядре клетки-хозяина. Репродукция включает раннюю и позднюю стадии, однако они разграничиваются нечётко.
Ранняя стадия репродукции герпесвирусов. В ранней стадии синтезируются «ранние белки», кодируемые проксимальной третью молекулы ДНК. Они проявляют регуляторные свойства, включая активацию транскрипции других участков вирусного генома, кодирующих ДНК-полимеразу и ДНК-связывающие белки.
Поздняя стадия репродукции герпесвирусов. В позднюю стадию вирусная ДНК-полимераза индуцирует репликацию материнской ДНК. В результате образуются молекулы ДНК дочерней популяции. Часть дочерней ДНК считывают клеточные полимеразы, что вызывает транскрипцию концевых генов, кодирующих структурные протеины (белки оболочки и гликопротеины шипов).
Сборка дочерних популяций герпесвирусов осуществляется в ядре, где капсидные белки окружают молекулы ДНК, формируя нуклеокапсиды. Финальная стадия морфогенеза герпесвирусов — формирование суперкапсида на внутренней поверхности ядерной мембраны. Зрелые дочерние популяции отпочковываются от модифицированной ядерной мембраны, транспортируются через цитоплазму и выделяются наружу.
Поксвирусы. Репродукция поксвирусов.
Поксвирусы обладают наиболее сложным репродуктивным циклом, при этом синтезируется более 100 различных белков, входящих в состав вирионов (большинство образует наружную оболочку). Репродукция поксвирусов характеризуется следующими особенностями.
• Транскрипция ДНК начинается до полной депротеинизации вируса, так как она полностью осуществляется вирусными полимеразами.
• Репликация происходит только в цитоплазме и полностью независима от клеточных полимераз, так как, в отличие от прочих вирусов, поксвирусы имеют собственную ДНК-зависимую РНК-полимеразу, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий.
В репродуктивном цикле выделяют три стадии— начальную, раннюю и позднюю.
Рис. 5-4. Репликативный цикл ДНК-содержащих вирусов (на примере репродукции вируса герпеса). После адсорбции (1) вирус проникает в клетку путём слияния с мембраной (2). Нуклеокапсид транспортируется к ядерной оболочке (3), и вирусная ДНК (вДНК) проникает в ядро клетки, где начинается её транскрипция клеточной ДНК-зависимой РНК-полимеразой (4). Вначале транскрибируются «ранние гены». В результате трансляции «ранней порции» вирусного генома синтезируются «ранние белки» (5), включая регуляторные, матричные и вирусные полимеразы. Вирусная полимераза проникает в геном клетки (6), где запускает синтез молекул ДНК дочерних популяций (7). Часть вирусной ДНК («поздние гены») дочерних популяций транскрибируется клеточной РНК-полимеразой (8), что приводит к синтезу «поздних белков» (9), необходимых для сборки дочерних популяций (10). Последние покидают ядро, отпочковываясь от его мембраны (11), фрагменты которой содержат в составе своей обопочки.
Начальная стадия репродукции поксвирусов запускается сразу же после раздевания вируса и выхода вирусной ДНК в цитоплазму.
Ранняя стадия репродукции поксвирусов. На этом этапе транскрибируется примерно половина вирусной ДНК. Синтезируются ферменты, кодируемые «ранними генами» и участвующие в репликации вирусной ДНК. Параллельно в небольшом количестве образуются структурные белки.
Поздняя стадия репродукции поксвирусов совпадает с началом репликации ДНК (что переключает механизмы транскрипции на считывание второй половины генома). Регуляторные белки блокируют трансляцию «ранней мРНК» и запускают синтез поздних (структурных) белков. Сборка вирионов осуществляется только в цитоплазме посредством реакций мембранного синтеза. Высвобождение зрелых популяций сопровождается лизисом клетки.
– Также рекомендуем “Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.”
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 19 февраля 2019;
проверки требуют 10 правок.
Герпесви́русы[3] (лат. Herpesviridae) — большое семейство ДНК-содержащих вирусов, вызывающее разнообразные болезни не только у человека и других млекопитающих, но и у птиц, рептилий, амфибий, рыб. Герпесвирусами болеют большинство населения нашей планеты[4].
Персистенция (вирусов) (лат. persistere — оставаться, упорствовать) — термин, предложенный в 1923 году французским бактериологом, иммунологом и вирусологом Константином Левадити (Levaditi Constantin, 1874—1953) и румынским вирусологом Штефаном Николау (Nicolai Stefan, 1896—1967), обозначающий длительное сохранение вируса в организме хозяина или в клеточной культуре.
Название семейства происходит от греч. ἕρπειν (herpein) ‘ползать’ через латинское herро ‘ползаю’. Инфекционные болезни, вызванные вирусами этого семейства, протекают остро, переходят в латентную стадию, не проявляют себя до определённого времени.
При ухудшении состояния организма (простуда, стресс, утомление и т. п.) вирус даёт о себе знать болезнью. Примером может служить герпетические высыпания на губах при «простуде».
Вирусы семейства Herpesviridae имеют общие биологические свойства. Они имеют эффективные механизмы взаимодействия с иммунной системой хозяина, позволяющие им достичь максимального распространения и сохраняться в организме в течение всей жизни.
В ходе своей жизнедеятельности вирусная ДНК экспрессирует определённые группы генов и, соответственно, кодируемых ими белков, которые, собственно, и определяют жизненный цикл вируса в клетках хозяина, приводя к изменению их фенотипических свойств, то есть трансформации.
Трансформация клеток вызывает развитие определённых иммунопатологических реакций, направленных против собственного организма и приводящих к вирусиндуцированной иммуносупрессии и длительной персистенции вируса в организме человека. В клетках своего хозяина они переходят в латентное состояние. При латентном состоянии нарушается полный репродуктивный цикл вируса. Он находится в клетках хозяина в виде субвирусных структур.
Клинические формы герпесвирусных инфекций характеризуются выраженным полиморфизмом. Существуют различные формы проявления герпесвирусных инфекции.
В любом случае герпесвирусы остаются в инфицированном организме пожизненно. Вирус может периодически рецидивировать с характерными клиническими проявлениями или бессимптомно, или приобретать генерализованный характер с возможным летальным исходом.
Вирионы семейства вирусов герпеса имеют сферическую форму диаметром от 120 до 300 нм. Внутри зрелой вирусной частицы содержится 35—45 различных белковых молекул.
В центре вириона различают сердцевину (кор, от лат. cor) размером 75 нм, содержащую ДНК.
Эта сердцевина окружена оболочкой — капсидом. Размер капсида составляет 100—110 нм. Он построен из множества одинаковых геометрически правильных белковых структур — капсомер. Капсомеры точно подобраны и подогнаны друг к другу, образуя двадцатигранник, по-гречески икосаэдр. Капсид содержит 162 капсомера: 150 шестиугольных (гексамеров) и 12 пятиугольных (пентамеров).
Вокруг капсида имеется аморфный белковый тегумент и всё это заключено в оболочку с гликопротеиновыми шипами.[источник не указан 2332 дня]
Геном герпесвирусов представляет собой линейную двухцепочечную молекулу ДНК. У герпесвируса человека 1-го типа она имеет размер 152 261 пара нуклеотидов и содержит 77 генов, все из которых кодируют белки; доля пар ГЦ составляет 68 %[5]. Геном герпесвируса человека 2-го типа состоит из 154 746 пар нуклеотидов и несёт 77 генов; доля пар ГЦ составляет 70 %[6].
Герпесвирусами заражаются человек и животные.
Образование новых вирусных частиц идёт под управлением вирусного генома. При попадании в организм носителя герпесвирусы адсорбируются клетками-мишенями на клеточной поверхности и освобождаются от капсида и дополнительной оболочки-конверта.
После эндоцитоза вирусной сердцевины происходит интегрирование вирусной ДНК с ядерным материалом клетки. Затем на ядерной мембране происходит образование и созревание новых вирионов и последующее их отпочковывание клеткой посредством экзоцитоза.
Для оболочки вируса, его капсида и ДНК, используются аминокислоты, белки, липопротеиды, нуклеозиды клетки-хозяина. По мере истощения внутриклеточных резервов, эти молекулы поступают в инфицированную клетку из межтканевых пространств.
Для культур, хронически зараженных вирусом простого герпеса, характерно[7]:
- возникновение состояния носительства после деструкции основной массы клеток и репопуляции немногих выживших;
- постоянное наличие очагов дегенерации, морфологически характерных для этого вируса;
- небольшой процент инфицированных клеток;
- низкая продукция вируса одной инфицированной клеткой;
- отсутствие интерферона в среде
- определенная резистентность клеток, освободившихся от состояния носительства, к реинфекции гомологичным вирусом.
В природе насчитывается восемь типов вирусов из семейства Herpesviridae, вызывающих болезнь у человека.
Семейство герпесвирусов по классификации Международного комитета по таксономии вирусов (ICTV) делится на подсемейства. Используются критерии:
- структура генома вируса
- белковый состав
- характер репликации вируса
- среда носительства
- тропизма к тканям
- распространение вируса в культуре
- длительности репродуктивного цикла
Под этим признаком различают подсемейства:
- Alphaherpesvirinae — альфагерпесвирусы, или α-герпесвирусы
- Betaherpesvirinae — бетагерпесвирусы, или β-герпесвирусы
- Gammaherpesvirinae — гаммагерпесвирусы, или γ-герпесвирусы
- 1 вид, не входящий ни в одно подсемейство и род (incertae sedis)
В 2016 году всем видам семейства изменили научное название для улучшения номенклатуры видов, при этом существенно не меняя названия герпесвирусов человека — к слову virus добавили префикс (alpha-, beta-, gamma-), указывающий подсемейство, к которому этот вид относится[8].
Выделяют восемь типов вирусов герпеса, вызывающих разные по тяжести процесса заболевания у людей. Характерной особенностью заболеваний является нахождение вирусов в организме человека в латентном состоянии[9][10].
- Human betaherpesvirus 6A
- Human betaherpesvirus 6B
- Human herpesvirus 6A, HHV-6A
- Human herpesvirus 6B, HHV-6B
- Богадельников И. В. Роль герпесвирусной инфекции в организме человека // Сучасні медичні технології. — 2009. — № 3. — С. 57-61.
Герпесвирусы (от греч. herpes— ползучий) —семейство крупных оболочечных ДНК-содержащих вирусов, вызывающих разнообразные инфекции.
Семейство Herpesviridae включает три подсемейства, отличающиеся по структуре генома, тканевому тропизму, цитопатологии и локализации латентной инфекции:
- подсемейство Alphaherpesvirinae — вирусы герпеса (ВПГ-1, ВПГ-2, VZV): для этой группы характерен быстрый рост. Вирусы размножаются в эпителиальных клетках, вызывая цитолитическое действие. В нейронах вызывают латентную, персистирующую инфекцию;
- подсемейство Betaherpesvirinae — вирусы герпеса (ЦМВ, ГВЧ-6, ГВЧ-7): для этой группы характерен медленный рост (латентная инфекция) в клетках эпителия слюнных желез, в гландах, почках, лимфоцитах. Вирусы оказывают цитомегалическое действие (ЦМВ) и лимфопролиферативное действие;
- подсемейство Gammaherpesvirinae — вирусы (ВЭБ) растут в лимфобластоидных клетках, оказывают лимфопролиферативное действие. Вызывают латентную инфекцию в лимфоидной ткани, лимфоцитах, эпителиальных клетках рта и глотки, слюнных желез. ВЭБ вызывает размножение В-лимфоцитов и персистирует в них.
Структура. Вирион герпесвируса имеет овальную форму диаметром 150-200 нм.
В центральной части вириона находится двунитевая линейная ДНК, окруженная икосаэдрическим капсидом, состоящим из 162 капсомеров. Снаружи вирус окружает оболочка с гликопротеиновыми шипами, сформированными из внутреннего слоя ядерной мембраны клетки. Пространство между капсидом и оболочкой называется тегумент (содержит вирусные белки и ферменты, необходимые для инициации репликации). Геном — двунитевая линейная ДНК. Она состоит: у ВПГ и ЦМВ — из двух фрагментов: короткого S и длинного L, каждый из которых у ВПГ заключен между двумя наборами инвертированных повторов, позволяющим геному рекомбинировать с образованием 4 изомеров; у VZV ДНК также состоит из двух фрагментов: короткого S и длинного L, но содержит один набор инвертированных повторов, поэтому формируется две изомерные формы.
После прикрепления к рецепторам клетки оболочка вириона сливается с клеточной мембраной (1, 2). Освободившийся нуклеокапсид (3) доставляет в ядро клетки ДНК вируса. Далее происходит транскрипция части вирусного генома (с помощью клеточной ДНК-зависимой РНК-полимеразы); образовавшиеся иРНК (4) проникают в цитоплазму где происходит синтез (трансляция) самых ранних альфа-белков (I), обладающих регулирующей активностью. Затем синтезируются ранние бета-белки (П) — ферменты, включая ДНК-зависимую ДНК-полимеразу и тимидинкиназу, участвующие в репликации геномной ДНК вируса. Поздние гамма- белки (Ш) являются структурными белками, включая капсид и гликопротеины (А, В, С, D, Е, F, G, X). Гликопротеины диффузно прилегают к ядерной оболочке (5). Формирующийся капсид (6) заполняется вирусной ДНК и почкуется через модифицированные мембраны ядерной оболочки (8). Перемещаясь через аппарат Гольджи, вирионы транспортируются через цитоплазму и выходят из клетки путем экзоцитоза (9) или лизиса клетки (10).
Вирус простого герпеса относится к семейству Herpesviridae, роду Simplexvirus. Вызывает простой герпес (herpes simplex), характеризующийся везикулезными высыпаниями на коже, слизистых оболочках, поражением центральной нервной системы и внутренних органов, а также пожизненным носительством (персистенцией) и рецидивами болезни. Вирус простого герпеса включает два типа: ВПГ-1 и ВПГ-2; распространен повсеместно, поражает большую часть населения Земли и существует в организме в латентной форме до момента реактивации.
ВПГ-1 поражает преимущественно область рта, глаз, ЦНС, а ВПГ-2 — гениталии, за что и получил название генитального штамма.
Читайте также: