Рост скелетных мышц. Регенерация скелетных мышц. Сухожилия.
Добавил пользователь Cypher Обновлено: 08.01.2026
1 ФГБОУ ВО «Пермский государственный медицинский университет им. академика Е.А. Вагнера» Минздрава России
В статье изложен обзор некоторых литературных данных о механизмах работы мышц во время различных по интенсивности нагрузок. Проведён анализ адаптивности, пластичности и степени изменения мышц на разных уровнях под влиянием различных факторов. Эти изменения связаны с увеличением мускульной силы и сопровождаются трансформацией одних типов мышечных волокон в другие, изменением метаболических процессов и увеличением синтеза сократительных белков, что в основном и обуславливает процессы мышечной гипертрофии. Главным пусковым механизмом данных процессов является синтез ростовых факторов, которые выполняют, по сути, основополагающую роль. Кроме усиления синтеза белка, данные факторы способны влиять и на миосателлиты. Именно эти клетки, располагающиеся на периферии симпласта, способны делиться в постнатальном периоде. В условиях наличия нагрузки и как следствия появления развивающихся микротравм происходит их активация. Они составляют основу регенерации поврежденного мышечного участка. За счёт своего деления они формируют новые мышечные волокна. Данный процесс очень напоминает воспалительный ввиду участия провоспалительных клеток. Их цитокины также влияют на пролиферацию. Поэтому при мышечной работе происходит множество процессов, которые способствуют к непосредственному увеличению мышечного волокна путём как за счёт гипертрофии, так и гиперплазии.
1. Волков Н.И., Несен Э.Н., Осипенко А.А., соавт. Биохимия мышечной деятельности // M.: «Олимпийская литература», 2000. С. 503.
3. Lin Y. et al. PGC-1? is associated with C2C12 Myoblast differentiation // Central European Journal of Biology. 2014. Т. 9. № 11. Р. 1030–1036.
4. Scharf M. et al. Mitogen-activated protein kinase-activated protein kinases 2 and 3 regulate SERCA2a expression and fiber type composition to modulate skeletal muscle and cardiomyocyte function // Molecular and cellular biology. 2013. Т. 33. № 13. Р. 2586–2602.
5. Meissner J.D. et al. The p38?/? mitogen-activated protein kinases mediate recruitment of CREB-binding protein to preserve fast myosin heavy chain IId/x gene activity in myotubes // Journal of Biological Chemistry. 2007. Т. 282. № 10. Р. 7265–7275.
6. Potthoff M.J., Olson E.N. MEF2: a central regulator of diverse developmental programs // Development. 2007. Т. 134. № 23. Р. 4131–4140.
На протяжении многих лет изучение процессов синтеза белков в скелетных мышцах при выполнении различных физических нагрузок остаётся актуальной проблемой биохимии и физиологии. Мышцы и их силовые характеристики очень важная составляющая организма каждого спортсмена, которая позволяет достигать результатов. В связи с прогрессивным развитием спорта и вовлечением большого количества людей в физическую культуру, тема здоровья спортсменов становится все более актуальной, интересной и увлекательной. Учитывая существующую сильную корреляцию между площадью поперечного сечения мышц и мышечной силой, стремление увеличить мышечную массу тела есть у каждого человека, занимающегося спортом. Кроме этого, необходимо помнить, что преобладание мышечной массы в организме благоприятно влияет на метаболические процессы.
Скелетная мышца – одна из наиболее пластичных структур в организме млекопитающих. При повышенной активности и нагрузке часто происходит увеличение её размеров, объёмов миофибриллярного аппарата, повышение сократительных возможностей (силы, мощности). Процесс прироста мышечной массы зависит от различных факторов: наследственных, конституциональных, а также пола, возраста, метаболизма, гормонального фона. Кроме того, с приобретением опыта тренировок становится все труднее увеличить мышечную массу, поэтому важно понимать и активно использовать все возможные механизмы этого процесса.
Клетки поперечно-полосатой мускулатуры отличаются от гладкомышечных миоцитов. Клетки скелетных мышц образуют многоядерный синцитий, основное вещество которого формируют миофибриллы, состоящие из толстых и тонких миофиламентов. Первый тип образуют молекулярные единицы и миозин, а второй тип содержит тропомиозин с тропонином и F-актин. Многие авторы считают скелетную мускулатуру гетерогенной системой относительно устройства и выполняемых функций, несмотря на её строгую организацию. Данное свойство помогает мышцам соответствовать возлагаемой на них функции. Так путём изменения количества саркомеров и миофибрилл обеспечивается их функциональная реорганизация [1].
Работа мышц проявляется их сокращением, которое начинается с появления очага возбуждения на нейромышечных окончаниях. Наружная мембрана деполяризуется, открываются кальциевые каналы, и концентрация кальция внутри клетки возрастает. Ионы кальция связываются с тропонином, при этом конформируется тропониновый комплекс. Участки цепей миозина связываются с актином, что сопровождается высвобождением энергии вследствие расщепления АТФ до АДФ и остатка фосфорной кислоты. Угол между лёгкой и тяжёлой цепями миозина изменяется и актиновый филамент перемещается к центру саркомера, что приводит к изменению длины мышцы, её сокращению [1, 2].
Клетки скелетных мышц подразделяются на два типа:
А) Миосателлиты – взрослые стволовые клетки мышечной ткани. Представляют собой основу для обновления мышц и прироста их массы;
Б) Миосимпласты – формируют многоядерный синцитий. Сами по себе являются мышечными тубами с миофибриллами внутри, по периферии которых располагаются ядра.
Нагрузки, оказываемые на мышцы, и само мышечное сокращение имеют некую зависимость. Предполагается, что первое будет напрямую соответствовать второму. Это достигается за счёт усиления экспрессии генов сократительных белков и энзимов обменных процессов. Мышечная активность сопровождается количественными и качественными изменениями в миоцитах того типа, которые необходимы для наиболее эффективного осуществления выполняемой работы [2].
Мышечные волокна делятся на медленные (I тип) и быстрые (II тип). Оба этих типа имеют различный состав, включающий в себя сократительные белки, ферменты энергетического обмена и внутриклеточный кальций.
Увеличение силы мышц проявляется структурными перестройками, которые затрагивают нервную и мышечные системы. Изменения в нервной системе проявляются трансформацией величины кортикальных полей, которые регулируют выполнение определённого вида движения, влиянием на синхронизацию моторных единиц и на обучение определенных мышц, отвечающих за выполнение данного вида движений. Таким образом, наибольшая активность мышц наблюдается именно тогда, когда она необходима для достижения максимального эффекта (активность мышц агонистов при одновременной пассивности антагонистов). Также наблюдается изменение частоты и устойчивости генерируемых импульсов и порога возбудимости мотонейронов. Изменения в мышечной системе могут быть связаны с гипертрофией скелетных мышц (увеличение размеров мышечного волокна) и с их гиперплазией (увеличение количества миоцитов) [3].
Но прежде чем переходить к последним двум процессам, необходимо разобраться с изменениями, происходящими в самих мышцах. В момент выполнения работы миоцит подвергается действию физических и гуморальных факторов (пассивные механические силы, гипоксемия, факторы роста, и т.д.). Они являются причиной запуска путей передачи сигнала внутри клеток, опосредуя транскрипцию и трансляцию генов, ответственных за синтез белков [2]. Изменения данных путей сопровождаются реорганизацией мышечных волокон, точнее их типов.
Одним из основных исходных сигналов является повышенная концентрация кальция внутри клетки и кальцинейрина. Кальцинейрин дефосфорилирует факторы транскрипции – NFAT (nuclear factor of activated T-cells), которые находятся в фосфорилированном состоянии [4]. Данные факторы в дефосфорилированной форме активируют гены-мишени, что способствует перестроению быстрых волокон в медленные.
По мере приспособления мышц к нагрузкам изменяются и процессы метаболизма в них. Существуют различные параметры, влияющие на формирование адаптивных механизмов в миоцитах при выполнении работы. Важнейшим является гипоксия, которая, в свою очередь активирует ферментные системы (фумараза, цитратсинтаза, ЛДГ) и запускает работу факторов транскрипции (PGC1). При недостатке кислорода происходит активация одной изоформы семейства гипоксия-индуцированных факторов (HIF; hypoxia inducible factor), которая проникает в ядро, связывается с определенным участком ДНК и активирует гены, отвечающие за гликолиз, потребление кислорода и ангиогенез, увеличивая данные процессы. Некоторые гормоны также способны влиять на экспрессию генов в мышечных клетках. Это такие гормоны, как инсулин, гормон роста, которые вместе с кортизолом запускают катаболические реакции в условиях метаболического и энергетического истощения [3].
Стоит напомнить, что мышцы не являются постоянными клетками, а заменяются в течение жизни. Пролиферация необходима для предотвращения апоптоза клеток (регулируемый процесс клеточной гибели) и поддержания массы скелетных мышц. Это осуществляется через динамический баланс между синтезом белков в мышцах и их распадом. Мышечная гипертрофия возникает тогда, когда синтез белков превышает их распад.
Что же наблюдается при гипертрофии и гиперплазии мышечного волокна? При растяжении и сокращении мышц происходит образование факторов роста IGF и MGF, которые могут действовать как паракринно, так и аутокринно. С одной стороны, их действие проявляется в увеличении синтеза сократительных белков мышечных волокон. Основным участником данного механизма является фосфорилированная PKB [5]. Её активация начинается с влияния на мышцу нагрузки, которая приводит к синтезу гена, запускающего путь IGF/PI3K. В ткани имеется несколько изоформ, некоторые из них (IGF-1 и MGF), взаимодействуя с рецепторами приводят к конформационным изменениям. Через фосфорилирование ряда рецепторов и происходит активация PKB, способствующая развитию анаболических реакций [6].
С другой же стороны, происходит усиление пролиферации миосателлитов, их митотическая активность приводит к формированию новых клеток, а также сопровождается слиянием их с имеющимися мышечными волокнами или даёт возможность формировать новые. Миосателлиты расположены между базальной мембраной и сарколеммой. Покоящиеся клетки активируются непосредственно травмированием мышцы и в ответ на это начинают активно делиться и соединяться с частями поврежденного волокна. Под влиянием тяжёлой изнурительной работы происходит также активация данных клеток из-за образования многочисленных микротравм мышечного волокна. Вследствие этого наблюдается явление подобное процессам, происходящим при воспалении. В зону повреждения активно мигрируют нейтрофилы и макрофаги, которые активируют синтез ранее упомянутых факторов роста, регулирующих пролиферацию и дифференцировку миосателлитов. Мышечная гипертрофия отличается от мышечной гиперплазии. При гипертрофии мышц, увеличиваются сократительные элементы, и межклеточный матрикс расширяется для поддержки роста. Гиперплазия приводит к увеличению количества мышечных волокон. Гипертрофия сократительных элементов может происходить путем добавления саркомеров либо последовательно или параллельно.
В отечественной литературе не утихают споры о патогенетических аспектах мышечного роста. Чаще всего гипертрофию скелетных мышц человека рассматривают как их долговременную адаптацию к физическим нагрузкам различной направленности. Но существует понятие о кратковременной гипертрофии скелетных мышц – то есть изменение объема мышцы в результате одной силовой тренировки. Спортсмены, выступающие в соревнованиях по бодибилдингу или бодифитнесу хорошо знают, что объем мышц можно немного увеличить за счет собственной крови и осмотического давления, если использовать специальный метод тренировки – пампинг.
Неоспоримым является факт увеличения объёма мышечных волокон. Это так называемая миофибриллярная гипертрофия, при которой происходит изменение объёма миофибрилл и плотность их укладки. Механизм связан с увеличением количества саркомеров в миофибриллах. Значительная роль при этом отводится активированным клеткам-сателлитам. Миогенные стволовые клетки начинают пролифелировать, а затем сливаются с существующими клетками или взаимодействуют между собой для формирования новых мышечных волокон. Этот механизм актуален при восстановлении травмированных клеток и при спортивной гипертрофии.
Существует множество данных, доказывающих идущий параллельно с этим процесс увеличения объёма несократительной части мышцы – саркоплазматическая гипертрофия. Это тонкие перестройки на биохимическом уровне клетки, а так же увеличение количества митохондрий. Многие авторы считают, что трансформации в саркоплазме повышают выносливость мышц. Ряд исследователей утверждает, что увеличение различных неконтрактильных элементов и жидкости действительно может привести к приросту мышечной массы, но без сопутствующего увеличения силы. Саркоплазматическая гипертрофия достигается специальными тренировками и часто описывается как нефункциональная. Однако ряд специалистов предполагают, что отек мышечных волокон вызывает увеличение синтеза белка и таким образом способствует росту сократительной ткани.
Эти процессы редко бывают сбалансированными и зависят от характера и интенсивности нагрузки. В скелетных мышцах при этом синтез мышечных белков преобладает над их распадом. Причиной такого метаболизма сторонники гипотезы ацидоза считают накопление молочной кислоты. С точки зрения другой теории – временная гипоксия запускает реперфузию мышц и активирует деление клеток-сателлитов. Последнее время широкое распространение получила гипотеза механического повреждения мышечных волокон. Микроразрывы сократительных белков и повреждения саркоплазмы сопровождается увеличением концентрации ионов кальция, что и стимулирует пролиферацию сателлитов.
Из этого следует, что механизмы мышечной гипертрофии известны и неоспоримы. Очень дискутабельным остается вопрос о наличии процесса гиперплазии мышц. Большинство авторов сходится во мнении, что увеличение количества мышечных волокон у человека не доказано, но при этом описывается возможность получения гиперплазии мышц в экспериментальных условиях у животных (млекопитающих и птиц). Некоторые исследователи допускают частичное увеличения числа волокон. На основании проведенного мета-анализа экспериментальных работ отмечено, что количество мышечных элементов увеличилось в экспериментах на птицах значительнее, чем при использовании в качестве подопытных млекопитающих. Примечательно также, что эффект гиперплазии наблюдался там, где использовались постоянные растяжения, а не упражнения, сочетающие его с расслаблением. Ряд исследователей (Kraemer, William J. и MacDougall J.) утверждают, что этот механизм может осуществляться под влиянием силовых тренировок. Однако доказательств увеличения мышечных волокон у людей недостаточно. Длительных исследований (более года) добровольцев и спортсменов не проводилось. Высказывается мнение, что это слишком короткий период для этого процесса. Гиперплазия подтверждается в биопсийном материале, а погрешность этого метода составляет около 10 %, что делает результат очень сомнительным.
Общее число волокон предопределяется генетически и практически не меняется в течение жизни без применения специальных стимуляторов. Российские ученые подтверждают, что вклад гиперплазии в процесс увеличения объема мышц составляет не более 5 % и, как правило, потенцирован использованием анаболических стероидов. Также гиперплазию могут вызывать блокаторы миостатина. Гормон роста при этом не вызывает гиперплазии.
Таким образом, при мышечной работе происходит множество процессов на разных уровнях. Начиная с изменений интенсивности обменных процессов и заканчивая изменениями механизмов нервной и гуморальной регуляции. Реорганизация мышц, лежащая в основе этих процессов, приводит к изменению многочисленных характеристик деятельности спортсменов.
Проанализировав все данные и изучив все возможные гипотезы, становится очевидным, что в увеличении мышечных волокон играют некую роль всё-таки два процесса. Первый – гипертрофия с ёе подвидами для сократительной и несократительной части мышцы (миофибриллярная и саркоплазматическая), которая, по мнению многих исследователей, занимает основополагающую роль. И второй это гиперплазия с её минимальным, но существенным вкладом.
Рост скелетных мышц. Регенерация скелетных мышц. Сухожилия.
Рост скелетных мышц. В период постнатального роста организма вместе с удлинением костей происходит рост в длину и толщину скелетных мышц. Масса скелетных мышц увеличивается также при интенсивных физических упражнениях. При этом рост мышц обусловлен утолщением (гипертрофией) мышечных волокон. Количество же их существенно не меняется. В основе гипертрофии мышечных волокон лежит увеличение числа миофибрилл и других органелл.
Новые миофибриллы образуются из миофиламентов, которые строятся в свою очередь из синтезируемых сократительных белков. Рост мышечных волокон в длину осуществляется в результате слияния с ними миосателлитоцитов. Удлинение миофибрилл достигается пристройкой к их концам новых саркомеров. Это происходит в местах прикрепления мышечных волокон к сухожилиям.

Регенерация скелетных мышц. После повреждения скелетных мышц часть мышечных волокон в области травмы подвергается разрушению. Продукты их распада резорбируются путем фагоцитоза и аутолиза. На месте травмы разрастается грануляционная ткань, в которую врастают мышечные элементы регенерата. Источником восстановления мышц являются в основном миоса-теллитоциты, способные делиться митозом и превращаться в миобласты. Последние сливаются друг с другом, формируя миосимпласты.
В ходе последующей их дифференцировки образуются новые мышечные трубочки и зрелые мышечные волокна. Процесс восстановления поврежденной мышцы продолжается около месяца. Для регенерации скелетной мышцы имеет важное значение восстановление иннервации (нервно-трофическое влияние), кровоснабжения и функционального натяжения. После тяжелых травм значительные дефекты скелетных мышц полностью не восстанавливаются. Обычно в таких случаях место мышечной травмы заполняется фиброзной рубцовой тканью. Этот процесс, как правило, сопровождает заживление мышечной раны после огнестрельного повреждения.
Сухожилия
С помощью сухожилий обеспечивается связь скелетных мышц с костями. Сухожилия образованы плотной оформленной соединительной тканью. В сухожилиях коллагеновые пучки находятся постоянно в состоянии натяжения. В процессе гистогенеза сухожилий при дифференцировке сухожильных клеток образуются коллагеновые волокна, располагающиеся параллельно друг другу. Каждый пучок коллагеновых волокон отделяется от соседнего слоем сухожильных клеток (тендоцитов). В целом этот комплекс клеток и волокон называется сухожильным пучком 1-го порядка.
Группа пучков 1-го порядка, окруженная прослойками рыхлой соединительной ткани, составляет сухожильный пучок 2-го порядка. Отделяющие эти пучки прослойки рыхлой соединительной ткани называют эндотенонием. Группа сухожильных пучков 2-го порядка образует сухожильный пучок 3-го порядка. Он окружен плотной соединительнотканной оболочкой — перитенонием. Несмотря на сильное развитие коллагеновых структур, оттесняющих на второй план клетки, сухожилия способны к регенерации. Камбиальным источником для восстановительных процессов служат малодифференцированные фибробласты и адвентициальные клетки в составе эндотенония.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Мышцы тела должны рассматриваться с точки зрения их развития и функции, а также топографии систем и групп, в которые они складываются.

Мышцы туловища развиваются из залегающей по бокам хорды и мозговой трубки дорсальной части мезодермы, которая разделяется на первичные сегменты, или сомиты. После выделения скелеротома, идущего на образование позвоночного столба, оставшаяся дорсомедиальная часть сомита образует миотом, клетки которого (миобласты) вытягиваются в продольном направлении, сливаются друг с другом и превращаются в дальнейшем в симпласты мышечных волокон. Часть миобластов дифференцируется в особые клетки — миосателлиты, лежащие рядом с симпластами. Миотомы разрастаются в вентральном направлении и разделяются на дорсальную и вентральную части. Из дорсальной части миотомов возникает спинная (дорсальная) мускулатура туловища, а из вентральной — мускулатура, расположенная на передней и боковой сторонах туловища и называемая вентральной (рис. 66, 67).
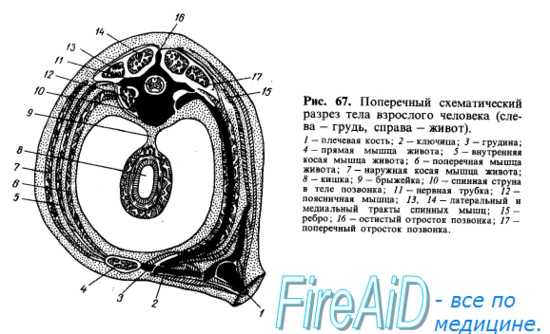
В каждый миотом (миомер) врастают ветви соименного спинномозгового нерва (невромера). Соответственно делению миотома на 2 части от нерва отходят 2 ветви, из которых дорсальная (задняя) входит в дорсальную часть миотома, а вентральная (передняя) — в вентральную. Все происходящие из одного и того же миотома мышцы снабжаются одним и тем же спинномозговым нервом. Соседние миотомы могут срастаться между собой, но каждый из сросшихся миотомов удерживает относящийся к нему нерв. Поэтому мышцы, происходящие из нескольких миотомов (например, прямая мышца живота), иннервируются несколькими нервами. Первоначально миотомы на каждой стороне отделяются друг от друга поперечными соединительнотканными перегородками, myosepta (рис. 68). Такое сегментированное расположение мускулатуры туловища у низших животных остается на всю жизнь. У высших же позвоноч.ых и у человека благодаря более значительной дифференцировке мышечных масс сегментация значительно сглаживается, хотя следы ее и остаются как в дорсальной (короткие мышцы перекидывающиеся между позвонками), так и в вентральной мускулатуре (межреберные мышцы и прямая мышца живота). Часть мышц, развившихся на туловище, остается на месте, образуя местную, аутохтонную мускулатуру (autos — тот же самый, chthon, греч. — земля). Другая часть в процессе развития перемещается с туловища на конечности. Такие мышцы называются трункофугальными (truncus — ствол, туловище, fugo — обращаю в бегство). Наконец, третья часть мышц, возникнув на конечностях, перемещается на туловище. Это трункопетальпые мышцы (рею — стремлюсь). На основании иннервации всегда можно отличить аутохтонную (т. е. развивающуюся в данном месте) мускула гуру от сместившихся в эту область других мышц-пришельцев
Мускулатура конечностей образуется из мезенхимы почек конечностей и получает свои нервы из передних ветвей спинномозговых нервов при посредстве плечевого и пояснично-крестцового сплетений. У низших рыб (селахий) из миотомов туловища вырастают мышечные почки, которые разделяются на два слоя, расположенные с дорсальной и вентральной сторон скелета плавника. Подобным же образом у наземных позвоночных мышцы по отношению к зачатку скелета конечности первоначально располагаются дорсально и вентрально (разгибатели и сгибатели). При дальнейшей дифференцировке зачатки мышц передней конечности разрастаются и в проксимальном направлении (трункопетальные мышцы) и покрывают аутохтонную мускулатуру туловища со стороны груди и спины (mm. pectorales major et minor, m. latissimus dorsi). Кроме этой первичной мускулатуры передней конечности, к поясу верхней конечности присоединяются еще трункофугальные мышцы, т. е. производные вентральной мускулатуры. служащие для передвижения и фиксации пояса и переместившиеся на него с головы (mm. trapezius и sternocleidomastoideus) и с туловища (mm. rhomboideus, levator scapulae, serratus anterior, subclavius, omohyoideus). У пояса задней конечности вторичных мышц не развивается, так как он неподвижно связан с позвоночным столбом. Сложная дифференцировка мышц конечностей наземных позвоночных, в особенности у высших форм, объясняется функцией конечностей, превратившихся в сложные рычаги, выполняющие различного рода движения.
Мышцы головы возникают отчасти из головных сомитов, а главным образом из мезодермы жаберных дуг. Висцеральный аппарат у низших рыб состоиг из сплошного мышечного слоя (общий сжиматель), который делится по своей иннервации на отдельные участки, совпадающие с мета-мерным расположением жаберных дуг: 1-й жаберной (мандибулярной) дуге соответствует V пара черепных нервов (тройничный нерв), 2-й жаберной (гиоидной) дуге — VII пара (лицевой нерв), 3-й жаберной дуге — IX пара (языког лоточный нерв). Остальная часть общего сжимателя снабжается ветвями X пары (блуждающий нерв). Сзади общего сжимателя обособляется пучок, прикрепляющийся к поясу верхней конечности (трапециевидная мышца). Когда с переходом из воды на сушу у низших позвоночных прекратилось жаберное дыхание- приспособленное для жизни в воде, мышцы жаберного аппарата (висцеральные) распространились на череп, где превратились в жевательные и мимические мышцы, но сохранили свою связь с теми частями скелета, которые возникли из жаберных дуг. Поэтому жевательные мышцы, возникающие из челюстной дуги и мышцы дна рта, располагаются и прикрепляются на нижней челюсти и иннервируются тройничным нервом (V пара). Из мускулатуры, соответствующей 2-й жаберной дуге, происходит главным образом подкожная мускулатура шеи и головы, иннер-вируемая лицевым нервом (VII пара).
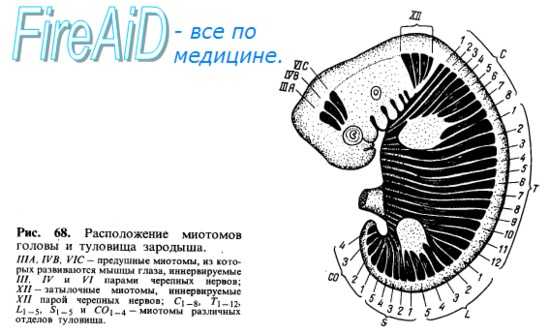
Мышцы, возникающие из материала обеих жаберных дуг, имеют двойное прикрепление и двойную иннервацию, например двубрюшная мышца, переднее брюшко которой прикрепляется к нижней челюсти (иннервация из тройничного нерва), а заднее — к подъязычной кости (иннервация из лицевого нерва). Висцеральная мускулатура, иннервируемая IX и X парами черепных нервов, у наземных позвоночных частью редуцируется, частью идет на образование мышц глотки и гортани. Трапециевидная мышца Теряет всякую связь с жаберными дугами и становится исключительно мышцей пояса верхней конечности. У млекопитающих от нее отщепляется в виде отдельной части грудино-ключично-сосцевидная мышца. Задняя ветвь блуждающего нерва, иннер-вирующая трапециевидную мышцу, превращается у высших позвоночных в самостоятельный черепной нерв — п. accessorius. Так как мозговой череп во всех своих частях представляет неподвижное образование, то на нем ожидать развития мышц нельзя. Поэтому на голове встречаются только некоторые остатки мускулатуры, образовавшейся из головных сомитов. К числу их нужно отнести мышцы глаза, происходящие из так называемых предушных миотомов (иннервация от III, IV и VI пар черепных нервов) (см. рис. 68).
Затылочные миотомы вместе с передними туловищными миотомами обычно образуют путем вентральных отростков особую поджаберную или подъязычную мускулатуру, лежащую под висцеральным скелетом. За счет этой мускулатуры, проникающей кпереди до нижней челюсти, происходят у наземных позвоночных мышцы языка, снабжаемые в силу своего происхождения из затылочных сомитов комплексом нервных волокон, образующих подъязычный нерв, который только у высших позвоночных стал настоящим черепным нервом. Остальная часть подъязычной мускулатуры (ниже подъязычной кости) представляет собой продолжение вентральной мускулатуры туловища, иннервируемой от передних ветвей спинномозговых нервов. Таким образом, для понимания расположения и фиксации мышц надо учитывать, кроме их функции, также и развитие (см. рис. 66, 67).
1 Медицинская академия им. С.И. Георгиевского, ФГАОУ ВО «Крымский федеральный университет им. В.И. Вернадского»
В статье представлены данные литературы о морфологических аспектах регенерации скелетных мышц. При повреждении скелетные мышцы подвергаются снижению функциональной активности, что является препятствующим фактором для выполнения той или иной мышечной активности. В работе представлены возможные варианты, которые могут уменьшить повреждение мышечного волокна. Известно, что скелетные мышцы. Скелетная мышца является симпластом, многоядерной структурой, которая образуется в онтогенезе вследствие слияния миобластов. Миоядра, или собственные ядра мышечного волокна, располагаются на периферии и в пространстве, которое находится между миофибриллами и сарколеммой. Также мышечное волокно содержит ядра, которые принадлежат фибробластам, эндотелиоцитам и миосателлитам. Скелетная мышечная ткань способна к регенерации после повреждения. Однако при большой потери мышечной массы для регенерации потребуется интервенционная поддержка. Следовательно, повреждение скелетной мышечной ткани является весомой проблемой в клинике. Для содействия восстановлению и регенерации мышц в течение последнего столетия и особенно в течение последних нескольких десятилетий были разработаны различные способы, в их числе хирургические методы, физиотерапия, использование биоматериалов, мышечная тканевая инженерия, а также клеточная терапия. Тем не менее, существует большая потребность в разработке новых методов и материалов, которые помогут восстановлению скелетных мышц и их функциональной регенерации.
2. Корсакова И. Н., Самчук Д. П., Еремин И. И., Зорин В. Л., Деев Р. В., Пулин А. А. Тканеинженерные конструкции для восстановления скелетной мышечной ткани. Гены и клетки. – 2017. doi: 10.23868/201703004
3. Курмашова Е.Д., Яковлева О.В., Ситдикова Г.Ф. Изменение процессов эндоцитоза синаптических везикул в условиях экспериментального сахарного диабета в различных типах мышц. Вестник науки Сибири. – 2015.
4. Шурыгин М. Г., Болбат А. В., Шурыгина И.А. Миосателлиты как источник регенерации мышечной ткани. Фундаментальные исследования. – 2015. doi: 611.018.6:616-003.93
6. Liu J., Saul D., Böker K.O., Ernst J., Lehman W., and Schilling, A.F. “Current Methods for Skeletal Muscle Tissue Repair and Regeneration,” BioMed Research International, 2018. doi: 10.1155/2018/1984879
8. M. V. Stevanovic, V. G. Cuéllar, A. Ghiassi, and F. Sharpe, “Single-stage Reconstruction of Elbow Flexion Associated with Massive Soft-Tissue Defect Using the Latissimus Dorsi Muscle Bipolar Rotational Transfer,” Plastic and Reconstructive Surgery - Global Open, 2016. doi:10.1097/gox.0000000000001066
10. Yiu E.M., Kornberg A.J., “Duchenne muscular dystrophy” , J Paediatr Child Health, 2015. doi: 10.1111/jpc.12868
Введение. Скелетная мышечная ткань является одной из самых распространённых в организме человека. На её долю приходится до 40-45% от общей массы тела. Существует порог регенерационного процесса скелетной мышцы, после превышения которого мышечная ткань теряет способность к восстановлению своих функций. Потеря скелетной мышечной ткани с длительным функциональным ухудшением определяется как «значительная потеря мышечной ткани» (VML - volumetric muscle loss), что может существенно повлиять на качество жизни человека, значительно снизив функциональность опорно-двигательного аппарата. Целью данной работы является анализ литературных источников с проведенными раннее исследованиями по восстановлению и регенерации тканей скелетных мышц.
Частыми причинами травм скелетных мышц являются дорожно-транспортные происшествия, травмы, полученные при взрывах, боевые ранения, хирургические и ортопедические манипуляции (например, после синдрома длительного сдавливания или резекции опухоли) или повреждения, случившиеся в ходе занятий спортом, приводящие к острой потере мышечной ткани. Повреждения, превышающие 20% мышечной массы, нуждаются в реконструктивных хирургических вмешательствах. Прогрессирующая мышечная потеря может быть следствием метаболических нарушений или наследственных генетических заболеваний, таких как мышечная дистрофия Дюшена, латеральный амиотрофический склероз и детская болезнь Шарко-Мари-Тута [10]. Атрофия мышц также может быть следствием травм периферических нервов, хронической болезни почек, сахарного диабета и сердечной недостаточности [3]. Потеря мышечной массы до 20% может быть компенсирована высокой адаптивностью и регенеративным потенциалом скелетных мышц. За этим порогом функциональные нарушения неизбежны и могут привести как к тяжелой инвалидности, так и к косметическим деформациям, поэтому терапевтическое вмешательство крайне востребовано для таких пациентов.
Анализ и обсуждение. Мышечная регенерация зависит от гетерогенной популяции сателлитных клеток, интерстициальных клеток и кровеносных сосудов и в основном контролируется с помощью белков межклеточного матрикса и секретируемых факторов [4]. В норме мышечная масса поддерживается балансом между синтезом и катаболизмом белка. В большинстве случаев VML тормозит регенерационную способность скелетных мышц, поскольку физически удаляются необходимые регенеративные элементы, в основном сателлитные клетки, периваскулярные стволовые клетки и базальная мембрана. Через денервацию активируются сигнальные пути распада белков (протеасомальные и аутофагиально-лизосомальные пути). Поэтому скорость разложения белка превышает его синтез, что способствует атрофии мышц, сопровождающейся постепенным снижением мышечной массы и диаметров мышечных волокон. Реваскуляризация обычно нарушается. Вследствие наступают ишемические условия способствующие пролиферации фибробластов, фиброзу и формированию фиброзной рубцовой ткани, что приводит к дальнейшей дистрофии мышцы. Состав и степень белков межклеточного матрикса в рубцовых тканях влияют на многие аспекты миогенеза, функции мышц и реиннервации. При хронической потере мышечной массы, такой как мышечная дистрофия Дюшена, фиброз является серьезной проблемой. Вследствие того, что последовательное разрушение миофибрилл не может быть полностью компенсировано пролиферацией сателлитных клеток, следующие воспалительные процессы приводят к изменению продукции белков внеклеточного матрикса и последующему развитию фиброза, а также формированию рубцовой ткани. Рубцовое образование можно уменьшить либо путем введения, например, 5-фторурацила и блеомицина, которые противодействуют пролиферации фибробластов и неоангиогенезу, либо с помощью лазерной терапии и функциональными улучшениями через 6-12 месяцев лечения [1]. Регенерация с регрессией рубцовой ткани и функциональным восстановлением может быть также оптимизирована с помощью трансплантации жира. Тем не менее, уменьшение фиброзирования недостаточно для восстановления и регенерации мышечного волокна. Клинические и научные исследования способствуют восстановлению большой мышечной потери. Современный стандарт лечения VML, как правило, основан на хирургическом вмешательстве с аутологичным мышечным трансплантатом и физиотерапией. Другими способами, которые применяют в клинике, являются: иглоукалывание и применение скаффолдов.
Хирургическое лечение VML включает в себя в основном санацию рубцовой ткани и / или транспозицию мышц. Аутологичная пересадка мышц обычно выполняется в клинической ситуации, когда после травмы, резекции опухоли или повреждения нерва возникают большие участки мышечной потери, что значительно ухудшает двигательную функцию. Хирурги трансплантируют здоровую мышцу из донорского участка, не затронутого повреждением, для восстановления утраченной или нарушенной функции. При отсутствии прилегающей мышцы из-за высокого уровня повреждений нерва или тяжелой травмы может быть применена аутологичная трансплантация мышцы в виде свободной функциональной пересадки. Часто используемыми аутологичными мышцами являются m. latissimus dorsi и m. gracilis. Было показано, что пересадка m. latissimus dorsi является безопасной и эффективной для восстановления сгибания локтевого сустава [8]. В случае синовиальной саркомы, поражающей m. gluteus medius и m. gluteus minimus, функция пораженного тазобедренного сустава может быть полностью восстановлена с помощью свободной нейроваскулярной трансплантации m. latissimus dorsi. Трансплантация m. gracilis обычно используется для восстановления локтевого сустава после травмы плечевого сплетения. Хотя данные мышечные лоскуты могут приводить к значительным терапевтическим результатам, они вызывают прогрессирующую заболеваемость донорского участка и недостаточность иннервации. Более того, до 10% этих реконструктивных операций приводят к полному отторжению трансплантата из-за таких осложнений, как инфекция и некроз.
Физические упражнения обладают способностью предотвращать снижение массы скелетных мышц. Таким образом, помимо хирургических методов, физиотерапия - это неинвазивный / минимально инвазивный способ улучшения восстановления и регенерации мышечной ткани. Данный метод получил широкое применение при восстановлении после травм и трансплантации мышечной ткани или для лечения хронической мышечной дистрофии. Физическая реабилитация направлена на укрепление сохранившихся мышц. Показано, что физиотерапия ускоряет заживление/регенерацию мышц, модулируя иммунный ответ, высвобождая факторы роста, способствуя васкуляризации и уменьшая образование рубцов. Физические упражнения и массаж, стимулирующие ангиогенез, являются потенциальными способами, ускоряющими образование новой мышечной ткани в клинически пересаженных мышечных трансплантатах или других хирургических операциях. Сообщалось, что физические тренировки могут регулировать сигнальный путь IGF-1 (Insulin-like growth factor-1) и уменьшать миостатин в мышечной ткани животных и людей, тем самым предотвращая атрофию мышцы. Физическая терапия действительно может улучшить восстановление мышц; однако этот метод не приносит весомых результатов при регенерации мышц у больных с VML. Кроме того, пациенты с тяжелыми заболеваниями или травмами часто не могут проводить последовательные упражнения, что ограничивает физическую терапию как лечение VML.
Акупунктура - это отрасль традиционной китайской медицины, которая широко используется для лечения различных заболеваний по всему миру. Было показано, что лечение электрическим иглоукалыванием подавляет экспрессию миостатина, приводя к пролиферации клеток-сателлитов и восстановлению скелетных мышц. Акупунктура + низкочастотная электрическая стимуляция (Acu-LFES - Acupuncture plus low-frequency electrical stimulation) усиливают регенерацию мышц и предотвращают потерю мышечной массы путем регулярных тренировок через стимуляцию сокращений. Такой метод лечения подходит для пациентов с тяжелыми заболеваниями, которые не могут часто выполнять упражнения. Было показано, что Acu-LFES противодействует атрофии скелетных мышц, вызванной диабетом, путем увеличения IGF-1 и тем самым стимулирует регенерацию мышц. Применение Acu-LFES для лечения диабетической миопатии и мышечной потери, вызванной хроническим заболеванием почек, показало хорошее функциональное восстановление мышц [9]. Основной механизм включает активацию макрофагов М2 и изменение уровней экспрессии мРНК E3 убиквитин-лигазы-атрогин-1. Подобно физическим упражнениям, иглоукалывание способствует восстановлению функции мышц и стимулирует их регенерацию, особенно у пациентов с атрофией мышц после хронических заболеваний. Тем не менее, не наблюдается значительного успеха при регенерации объемных мышечных дефектов после травмы или резекции опухоли. Кроме того, необходимо много времени, чтобы определить оптимальную продолжительность и интенсивность Acu-LFES в качестве стандартного лечения атрофии мышц.
Биологические скаффолды, состоящие из белков внеклеточного матрикса (ECM - the extracellular matrix), широко используются в регенеративной медицине и в хирургических процедурах для восстановления и регенерации тканей. Скаффолды могут способствовать восстановлению VML, обеспечивая структурную и биохимическую основу. Несколько тканевых скаффолдов были протестированы на животных моделях при незначительных потерях мышечной массы, впоследствии дали хороший результат и были апробированы в клинике. Ксеногенный внеклеточный матрикс и аутологичная ткань были использованы для восстановления функций мышц и одновременного формирования биологической ниши для регенерации. Многослойный каркас из белков межклеточного матрикса, полученный из подслизистой оболочки свиньи, был применен для восстановления у пациентов m. vastus medialis. Через 4 месяца после операции наблюдался заметный прирост показателей функциональной активности, а на месте имплантации была обнаружена новая мышечная ткань. Подслизистая оболочка тонкого кишечника свиньи также использовалась для лечения дефектов брюшной стенки [6]. Кроме того, белки межклеточного матрикса свиньи из мочевого пузыря были имплантированы в попытке лечения VML у человека. Функциональное улучшение с формированием мышечной ткани наблюдалось у трех из пяти пациентов в данном исследовании. Однако ксеногенные скаффолды все еще могут вызывать неблагоприятный иммунный ответ после децеллюляризации, и могут повысить риск передачи инфекционных заболеваний. Поэтому существует клиническая необходимость в разработке новых методов, которые смогут более эффективно восстановить поврежденную мышечную ткань.
Заключение. Повреждение скелетной мышечной ткани или атрофия мышц встречается довольно часто. Хирургические методы достигли значительных результатов и могут гарантировать хорошие результаты для восстановления мышечных функций. Однако, при хирургическом вмешательстве всегда остается риск, обусловленный как человеческим фактором, так особенностями операции. Исследования в области тканевой инженерии и регенеративной клеточной терапии могут решить данную проблему. Тканевая инженерия использует биологические скаффолды, которые направляют развитие мышечной ткани при участии факторов роста. Эти клетки способствуют пролиферации миогенных клеток в поврежденных или атрофических мышцах, что отражается на их дальнейшей регенерации. Подобные творческие подходы опираются на глубокое понимание процесса, необходимого для функционального восстановления мышц (реакция клеток на скаффолды, васкуляризация, миогенез и иннервация), однако все равно нуждаются в дальнейших исследованиях.
Адаптация скелетных мышц человека к физическим нагрузкам
Даны определения адаптации, стресса (общего адаптационного синдрома) и суперкомпенсации. Рассмотрены виды адаптации (срочная и долговременная) и условия адаптации скелетных мышц человека к физическим нагрузкам.
Организм человека устроен таким образом, что, попадая в новые для него условия, он может к ним приспособиться. Такое свойство организма человека получило название адаптация.
Понятия адаптации и стресса
Адаптация как общее универсальное свойство живого обеспечивает жизнеспособность организма в изменяющихся условиях и представляет собой процесс адекватного приспособления его функциональных и структурных элементов к окружающей среде. При этом основная задача адаптации состоит в поддержании постоянства внутренней среды организма – гомеостаза[1]. Термин «адаптация» тесно связан с понятием «стресс».
Стресс – неспецифическая (общая) реакция организма на воздействие, нарушающее его гомеостаз.
Г. Селье установил, что на разные по качеству, но сильные раздражители (стресс) организм для выравнивания гомеостаза всегда отвечает однотипными реакциями, которые были названы им общим адаптационным синдромом.
Спортивную тренировку можно рассматривать как адаптацию организма спортсмена к нагрузкам определенной направленности. При этом под воздействием систематических тренировочных нагрузок внутренняя среда организма человека претерпевает значительные изменения. После прекращения нагрузки в организме начинаются процессы, направленные на восстановление исходного состояния.
Виды адаптации
Различают срочную и долговременную адаптацию организма спортсмена к тренировочным воздействиям. Так как основным объектом этой статьи являются скелетные мышцы, вопросы адаптации будут рассматриваться в этом ракурсе.
Срочная адаптация – это структурно-функциональная перестройка, происходящая в организме спортсмена непосредственно во время выполнения физических упражнений. Основной целью срочной адаптации является создание оптимальных условий для функционирования мышц, прежде всего, за счет увеличения их энергоснабжения. В связи с этим, значительно ускоряются реакции катаболизма[2] при одновременном снижении скорости анаболических[3] процессов (в основном, синтеза белков), так как в ходе катаболических реакций выделяется энергия.
Долговременная адаптация – структурно-функциональная перестройка, происходящая в организме в ответ на длительное или многократное воздействие физической нагрузки. Долговременная адаптация протекает в организме спортсмена в промежутках между тренировками.
Условия адаптации
Первым условием является многократное (повторное) применение физических нагрузок. Однократная физическая нагрузка не вызывает стойких адаптационных перестроек в организме. Если же физические нагрузки повторяются, в организме создается необходимый метаболический фон, который обеспечивает постепенность формирования морфологических, биохимических и функциональных изменений. При повторяющихся тренировочных нагрузках благодаря активации генетического аппарата мышечных волокон в мышцах увеличивается содержание структурных и сократительных белков, вследствие чего мышцы становятся более резистентными к задаваемой нагрузке.
Вторым условием, определяющим процесс адаптации организма к физическим нагрузкам, является их регулярное применение Необходимость регулярно выполнять физические упражнения связана с изменениями метаболизма, которые происходят в организме в процессе физических нагрузок. В зависимости от интенсивности и длительности физической нагрузки, в организме происходят изменения в обмене веществ, которые могут быть ограничены локальными сдвигами в энергетическом обмене или затрагивать метаболизм всего организма. В последнем случае процесс восстановления метаболизма до уровня покоя занимает значительно больше времени и требует большего периода отдыха.
Долговременная адаптация организма к различным факторам внешней среды возможна благодаря явлению суперкомпенсации (сверхвосстановления). В области мышечной деятельности это явление первыми описали Л.И. Ямпольский (1949) и Н.Н. Яковлев (1949,1955). В основе суперкомпенсации лежит взаимодействие между нагрузкой и восстановлением. Это – циклический процесс и его пусковым стимулом является физическая нагрузка. После нагрузки возникает утомление и резкое снижение работоспособности спортсмена, что соответствует первой фазе цикла. Во второй фазе начинается процесс восстановления работоспособности. К концу второй фазы работоспособность спортсменов достигает исходного уровня. В третьей фазе (фазе суперкомпенсации) работоспособность превышает исходный уровень. Последняя, четвертая фаза характеризуется возвращением работоспособности к исходному уровню (рис.1 а).
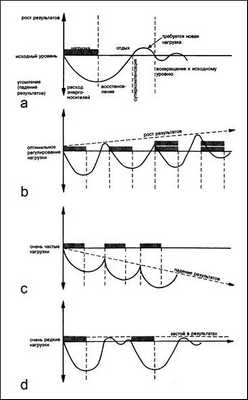
Рис.1. Варианты долговременной адаптации организма человека к физическим нагрузкам
Очень наглядно эффект суперкомпенсации виден на примере восстановления энергетических запасов мышц. Перед началом тренировки в мышцах находится определенное количество энергетических веществ (например, креатинфосфата, гликогена и др.). В результате тренировки происходит снижение уровня этих веществ в скелетных мышцах. После окончания тренировки, в фазе восстановления уровень энергетических веществ в мышечном волокне превышает исходный, то есть происходит суперкомпенсация.
Долговременная адаптация возможна только в том случае, если достигаемые срочный и отставленный тренировочные эффекты от каждой тренировки будут суммироваться (рис.1 b). Поэтому для получения определенного тренировочного эффекта и последующего повышения физической работоспособности очередную физическую нагрузку следует проводить в период преимущественно суперкомпенсации после предшествующей работы. Слишком частые (рис.1 c) тренировки прерывают стадию восстановления до достижения эффекта суперкомпенсации. Вследствие этого возможно постепенное развитие процесса недовосстановления работоспособности и преждевременное наступление утомления, что негативно сказывается на результатах. Слишком редкие тренировки (рис.1 d) в фазе сниженной суперкомпенсации не позволяют закрепить тренировочный эффект, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню.
Однако еще в начале ХХ века М.Е. Маршак (1931) установил, что процессы восстановления после тяжелой мышечной работы в различных вегетативных системах протекают гетерохронно, то есть с разной скоростью. Более того, даже в пределах одной и той же системы для разных показателей ее функции возвращаются к уровню покоя не одновременно. В последующем эти данные были подтверждены для многих физиологических и биохимических показателей.
Следует отметить, что процессы восстановления энергетических веществ в мышцах также протекают с разной скоростью и завершаются в разное время. Вначале из скелетных мышц и крови удаляется молочная кислота, которая окисляется до СО2 или включается в синтез гликогена, затем происходит ресинтез креатинфосфата, гликогена и жиров.
Существует правило Энгельгардта, согласно которому интенсивность протекания восстановительных процессов и сроки восстановления энергетических запасов организма зависят от интенсивности их расходования во время выполнения упражнения. Следовательно, чем больше расход энергетических запасов мышц при работе, тем интенсивнее идет их восстановление и тем значительнее превышение исходного уровня в фазе суперкомпенсации. Однако это правило применимо лишь в ограниченных пределах. На основании эмпирических исследований установлено, что во взаимоотношениях тренировочных раздражителей и адаптационных реакций лежат следующие закономерности (Ю. Хартманн, Х. Тюнеманн, 1988):
- подпороговые раздражители (30% нагрузки от максимально возможной) не вызывают никаких адаптационных сдвигов;
- слишком высокие раздражители (большой объем и интенсивность нагрузки) вызывают спад результатов;
- оптимальные нагрузки приводят к оптимальным результатам.
В процессы адаптации, возникающие при интенсивных физических нагрузках, вовлекаются все системы, обеспечивающие функционирование мышц. Наряду с увеличением синтеза сократительных белков мышц возрастает интенсивность функционирования систем, обеспечивающих их катаболизм.
В экспериментах на животных, проведенных П.З. Гудзем (1963), в которых животные вначале подвергались различным по длительности и интенсивности физическим нагрузкам, а затем помещались в тесные клетки, было установлено следующее. Если животные получали умеренные динамические нагрузки или повышенные статические, а затем были переведены в режим гиподинамии, длительное пребывание в тесных клетках не вызвало существенных деструктивных изменений в скелетных мышцах. Однако у животных, которые до этого получали повышенные физические нагрузки, через четыре недели пребывания в тесных клетках были обнаружены дистрофические и деструктивные изменения гипертрофированных мышечных волокон. Из этого можно сделать вывод, что адаптированные к высоким физическим нагрузкам системы, отвечающие за катаболизм белка, после резкого снижения физических нагрузок продолжают функционировать некоторое время с максимальной мощностью, что приводит к резкой дистрофии и деструктивным изменениям в мышечных волокнах.
Третьим условием достижения прочных адаптационных сдвигов является постепенное увеличение физических нагрузок как по объему, так и по интенсивности. Если это условие не будет соблюдено, то по мере адаптации организма к тренировочным нагрузкам будет постепенно снижаться величина энерготрат и изменения метаболизма будут менее выраженными (М.И. Калинский, В.А. Рогозкин, 1989).
С биохимической точки зрения можно выделить несколько факторов, изменения которых существенно влияют на обмен веществ тренированного с помощью физических нагрузок организма. Во-первых, в скелетных мышцах и других органах и тканях повышаются запасы энергетических ресурсов (креатинфосфата и гликогена). Во-вторых, расширяются потенциальные возможности ферментного аппарата: повышается активность ферментов гликолиза, цикла лимонной кислоты, окисления жирных кислот, систем транспорта ионов. В-третьих, улучшаются механизмы регуляции обмена веществ с участием нервной и эндокринной систем, а также внутриклеточной системы автономного регулирования. Все эти факторы – наличие повышенного количества энергетических ресурсов и увеличенная активность ферментных комплексов, обеспечивающих основные циклы энергетического метаболизма – открывают возможности для более быстрого и более длительного пополнения запасов АТФ в организме.
В отличие от креатинфосфата и гликогена, концентрация АТФ в тканях тренированного организма не возрастает, однако, меняется скорость обмена молекул АТФ, так как повышается каталитическая активность ферментов, участвующих в гидролизе АТФ во время мышечного сокращения и в процессе ресинтеза. Под влиянием физических нагрузок в скелетных мышцах увеличивается концентрация креатинфосфата и повышается активность фермента креатинкиназы, участвующего в ресинтезе АТФ. Это приводит к расширению энергетических ресурсов в мышце и повышению скорости восстановления АТФ из креатинфосфата.
Саркоплазматическая и миофибриллярная гипертрофии скелетных мышц возможны только в том случае, если будут соблюдены описанные выше условия.
- Маршак, М.Е. О восстановительном периоде после мышечной работы / М.Е. Маршак // Физиологический журнал СССР, 1931. – Т.14. – № 2-3. – С. 204.
- Яковлев Н. Н. Очерки по биохимии спорта. М.: Физкультура и спорт, 1955. 264 с.
- Ямпольский, Л.И. Расходование и ресинтез гликогена мышц в зависимости от характера мышечной деятельности / Л.И. Ямпольский: Автореф. дис…канд. биол. наук, 1949. – 20 с.
- Хартманн, Ю. Современная силовая тренировка / Ю. Хартманн, Х. Тюнеманн. – Берлин: Шпортферлаг, 1988. – 335 с.
- Самсонова А. В. Гипертрофия скелетных мышц человека: Учеб. пособие. СПб: Кинетика, 2018. 159 с.
[1] Гомеостаз» – относительное, колеблющееся в строго очерченных границах постоянство внутренней среды – крови, лимфы, тканевой и внеклеточной жидкости и устойчивость основных физиологических функций: артериального давления, ЧСС и др.
[2] Катаболизм – совокупность химических реакций, за счет которых крупные молекулы превращаются в молекулы меньшего размера.
Читайте также: