Сходства археев прокариот с эукариотами
Добавил пользователь Alex Обновлено: 08.01.2026
Организмы одноклеточных и многоклеточных делятся на две категории — эукариоты и прокариоты.
Клетки животных, а также почти все растения и грибы обладают интерфазным ядром. Кроме того, прокариотические и эукариотические клетки (прокариоты и эукариоты) имеют стандартные для всех клеток органоиды. Такие организмы называются ядерными или эукариотами.
Прокариоты или доядерные — это не такая большая категория организмов, как эукариоты, но более древняя по своему происхождению. К ним относятся бактерии сине-зеленые водоросли (цианобактерии). У них нет настоящего ядра и большинства органоидов, присущих цитоплазме.
Но у эукариот и прокариот есть свои особенности. Обратимся к сравнению клеток прокариот и эукариот, в частности, рассмотрим строение прокариотической и эукариотической клеток, а также обозначим различия прокариот и эукариот.
Сравнительная характеристика прокариот и эукариот
Характеристика клеток прокариот
При сравнении прокариот и эукариот важно подробно остановиться на строении.
Прокариотическая и эукариотическая клетки имеют разное строение. Строение клеток прокариот достаточно простое. Клетка прокариот не имеет ядра, ядрышка и хромосом. Клеточное ядро в этом случае заменяет нуклеоид. Он представляет собой похожее на ядро образование, без оболочки с одной кольцевой молекулой ДНК, которая связана с небольшим количеством белка. Также можно сказать, что это скопление белков и нуклеиновых кислот: они лежат в цитоплазме и не отделены от нее мембранами.
Последний момент является ключевым для деления клеток на прокариот и эукариот (доядерные и ядерные). Далее мы посмотрим сравнение эукариотических и прокариотических клеток в таблице.
В прокариотических клетках нет внутренних мембран — за исключением вмятин плазмолеммы. Исходя из этого получается, что органеллы прокариот немногочисленны: митохондрий, эндоплазматической сети, хлоропластов, лизосом, комплекса Гольджи. Все перечисленное есть в эукариотических клетках — там они окружены мембраной. Вакуоли также отсутствуют.
В прокариотических клетках есть только одна единственная органелла — это рибосома. Но здесь рибосомы мельче, чем у клеток эукариот.
Строение клетки прокариот характеризуется тем, что у клеток есть плотная клеточная стенка, которая их покрывает, и часто слизистая капсула.
Клеточная стенка состоит из муреина. Молекула муреина, в свою очередь, включает параллельно расположенные полисахаридные цепи: они сшиты друг с другом короткими цепями пептидов.
Плазматическая мембрана характеризуется тем, что у нее есть способность прогибаться внутрь цитоплазмы и образовывать, таким образом, мезосомы. На мембранах мезосом находятся окислительно-восстановительные ферменты, а фотосминтезирующие прокариоты имеют также соответствующие пигменты: бактериохлорофилл (бактерии) и фикобилины (цианобактерии). За счет этого мембраны получают возможность осуществлять функции, свойственные митохондриям, хлоропластам и другим органеллам.
Для прокариот характерно бесполое размножение. Оно происходит в результате простого деления клетки пополам.
Сравнительная характеристика клеток, представленных в таблице, поможет различать два типа клеток без каких-либо проблем.
Сравнительная характеристика прокариот и эукариот в таблице:
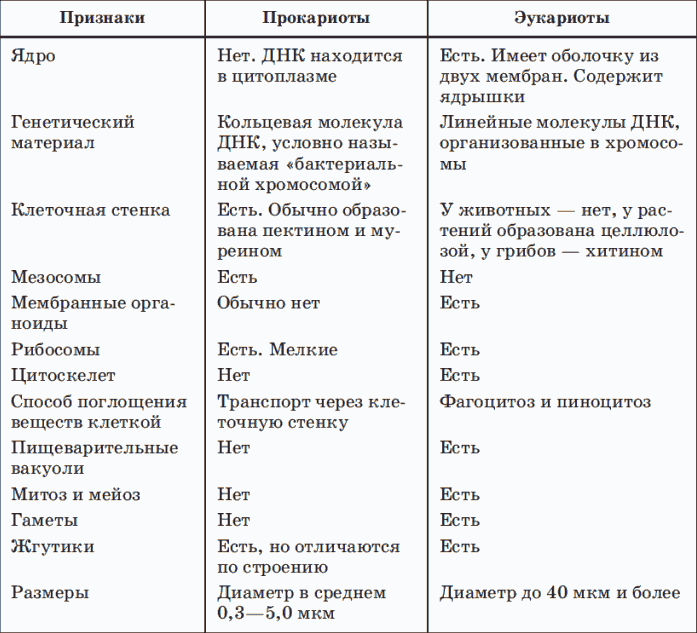
Если посмотреть на сравнение клеток прокариот и эукариот в таблице, то становится понятно, в чем заключается их похожесть и отличия. В таблице прокариоты и эукариоты — это практически две разные клетки.
Кстати, сравнение клеток прокариот и эукариот в таблице в 9 классе уже необходимо уметь делать.
Сравнительная характеристика эукариот и прокариот будет неполной без анализа первых. Так что помимо сравнительной характеристики клеток в таблице нужно знать, что собой представляют эукариоты.
Характеристика клеток эукариот
Эукариотическая и прокариотическая клетки обладают разным составом.
Несмотря на то, что клетки эукариот включают те же структурные элементы, что и прокариотические клетки, строение клетки эукариот сложнее. К таким элементам относятся цитоплазма, клеточная стенка эукариот, плазмолемма.
Строение клеток эукариот характеризуется разделением на компартменты (реакционные пространства) при помощи множества мембран. В каждом из компартментов происходят разнообразные химические реакции — одновременно и независимо друг от друга.
Ниже представлены сведения об эукариотической клетке в таблице (сравнение клеток разных царств эукариот не приводим).
Строение эукариотической клетки в таблице, а точнее, в одной картинке:
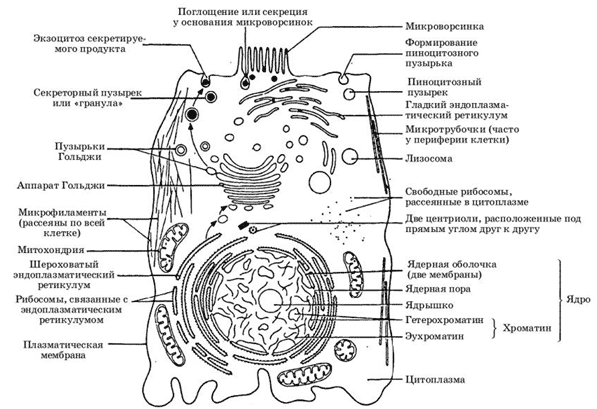
Из таблицы строения эукариотической клетки понятно, насколько сложным оно является.
Главные функции в клетке выполняют ядро и различные органеллы, такие как митохондрии, комплекс Гольджи, рибосомы и др. Что касается ядра, пластид и митохондрий, то они отделены от цитоплазмы при помощи двухмембранной оболочки. Генетический материал содержится в ядре клетки.
Функция хлоропластов — улавливание солнечной энергии и преобразование ее в химическую энергию углеводов при помощи фотосинтеза.
Митохондрии получают энергию в процессе расщепления белков, углеводов, жиров и других органических соединений.
Эндоплазматическая сеть и комплекс Гольджи — это мембранные системы цитоплазмы эукариотических клеток. Их наличие обеспечивает нормальное осуществление всех жизненных процессов в клетке.
Лизосомы, вакуоли и пероксисомы отвечают за выполнение специфических функций.
Немембранное происхождение имеют хромосомы, рибосомы, микротрубочки и микрофиламенты.
Основной способ размножения эукариотических клеток — митоз.
Эта основная информация по сравнению прокариотической и эукариотической клетки. Отличия прокариот от эукариот в таблице наглядно видны.
Сходства археев прокариот с эукариотами
• Археи стремятся адаптироваться к экстремальным условиям существования и использовать «необычные» источники энергии
• В составе клеточной оболочки археев находятся уникальные компоненты и отсутствует пептидогликан клеточной стенки
• По характеру основных процессов метаболизма и наличию некоторых структур, например жгутиков, археи напоминают бактерии
• По характеру репликации ДНК, транскрипции и трансляции археи напоминают клетки эукариот, однако в регуляции активности генов у них участвует много белков, напоминающих бактериальные
Прокариоты подразделяются на две группы, в настоящее время известные под названиями бактерии и археи. Вначале оказалось, что различия между этими группами проявляются в последовательности рРНК, однако они также отмечаются при сопоставлении их важнейших физиологических и биохимических характеристик. Особенно у археев, как следует из рисунка ниже. проявляется ряд общих свойств с клетками эукариот. Более того, хотя археи, несомненно, относятся к прокариотам, поскольку не обладают ядерной мембраной, они отличаются от хорошо известных бактерий по своим фундаментальным свойствам. В особенности это проявляется в отсутствии пептидогликана в их клеточной стенке и наличии в мембранах липидов, связанных с глицерином эфирной связью. Вообще говоря, об архе-ях мы знаем гораздо меньше, чем о бактериях.
Большая часть хорошо изученных видов археев характеризуется универсальностью метаболизма и способностью существовать в экстремальных условиях. Некоторые из них могут расти при очень высоких температурах (> 80 °С). Очевидно, что лишь один белок, обратная гираза, является специфичным для гипертермофильных археев. Таким образом, способность перекручивать ДНК может представлять собой единственный критический фактор, необходимый для адаптации этих микроорганизмов к условиям высокой температуры. Многие виды археев приспособлены к существованию в условиях экстремальных величин pH (ацидофилы и алкалофилы) или концентрации солей (галофилы).
К организмам, обладающим особенностями метаболизма, относятся метаногены (например, Methanococcus janaschii), которые в анаэробных условиях используют двуокись углерода, соединения содержащие метальную группу или ацетат; термофильные восстановители серы (например, Archaeglobus fulgidis) которые в качестве акцепторов электронов при дыхании используют окисленные соединения серы; и галофилы (например, Halobacterium salinarum), которые адаптировались к высокой концентрации соли. Crenarchaeotes (например, Sulfolobus solfataricus) представляют собой совершенно самостоятельную ветвь археев. Наиболее изученными представителями этой ветви являются термофилы, зависимые от источников серы. Филогенетическое родство основных групп археев представлено на рисунке ниже.
В настоящее время полностью отсеквенирован геном многих представителей археев. Так же как бактерии, археи имеют относительно компактные кольцевые хромосомы размерами порядка 1,5-3 мнп. Секвенирование генома позволило рассчитать способность каждого организма к кодированию белков. Исходя из рассчитанного набора белков, можно прийти к выводам относительно структуры и функций археев. В целом, больше всего сходства между археями и бактериями проявляется в клеточной структуре и в организации генома. У архей и бактерий существует много таких общих компонентов мембран и клеточной оболочки, как АВС транспортеры и капсулярные полисахариды. Археи также напоминают бактерии по характеру основных метаболических процессов и по наличию таких адаптивных функций, как подвижность, обусловленная наличием жгутиков, и хемотаксис.
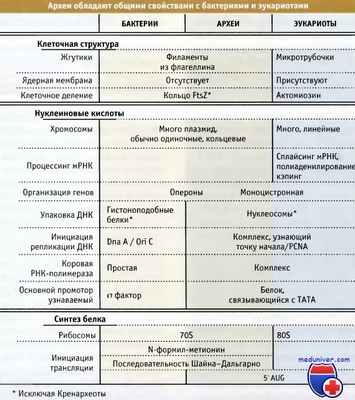
Археи обладают общими свойствами с бактериями и эукариотами.
Наряду с этим, в геноме археев содержится много инсерционных последовательностей и экстрахромосомальных элементов (например, плазмид), которые напоминают находящиеся в геноме бактерий.
Прочие системы археев обладают чертами, близкими к таковым систем бактерий и эукариот. Это, вероятно, относится к системе секреции белков и к белковым шаперонам. Большинство рибосомальных субъединиц археев обнаруживаются у бактерий и у эукариот, однако несколько субъединиц присутствуют только у эукариот и отсутствуют у бактерий. Факторы инициации трансляции и элонгации также напоминают факторы эукариот, а не бактерий. Однако некоторые аспекты инициации трансляции у археев больше похожи на бактериальные. У бактерий (и в митохондриях, и хлоропластах) перед стартовым кодоном находится последовательность «рибосомального сайта связывания» (последовательность Шайна-Дальгарно). Эта последовательность позволяет генам организоваться в опероны с несколькими стартовыми сайтами в одной мРНК. В цитоплазме эукариот узнавание сайта инициации трансляции связано с функционированием механизма отслеживания, при котором выбирается кодон АУГ, ближайший к 5'-концу мРНК.
Вероятно, этот механизм не позволяет транслировать более одного гена на молекулу мРНК. У археев, большинство генов имеет моноцистронную природу, и трансляция, вероятно, инициируется по тому же механизму, что и у эукариот. Однако у археев также есть опероны, где «ниже расположенным генам» предшествует последовательность рибосомального сайта связывания, характерная для бактерий.
Говоря об организации ДНК, отметим, что большинство археев содержат легко узнаваемые гомологи гистонов эукариот. Это согласуется с характером организации хромосом при участии нуклеосомоподобных структур, которые отсутствуют у бактерий. Crenarchaeota представляют собой исключение, поскольку у них не обнаружено ни гистонов, ни «гистоноподобных» белков бактерий. Репликация ДНК у археев происходит с участием полимераз, которые, по-видимому, напоминают соответствующие ферменты эукариот. Аналогичным образом, факторы инициации репликации ДНК и процессивность полимеразы у археев характерны для эукариот, а не для бактерий.
Аппарат транскрипции у археев достаточно сложен, и основная РНК-полимераза и факторы инициации транскрипции напоминают таковые у эукариот. Так, основная РНК полимераза обычно состоит из 11 субъединиц, большинство которых у бактериальных полимераз отсутствует. Инициация транскрипции контролируется факторами, в основном гомологичными соответствующим факторам у эукариот. Основным фактором, участвующим в распознавании базального промотора, является белок, связывающийся с последовательностью ТАТА (ТВР). У археев отсутствует сигма-фактор, являющийся ключевым участником в инициации транскрипции и в узнавании промотора у бактерий. В настоящее время мы еще мало знаем о регуляции транскрипции у археев.
Однако удивительно, что последовательности генома археев, вероятно, содержат много регуляторов транскрипции, близких к таковым бактерий. Это позволяет предполагать, что активация и репрессия генов у археев могут осуществляться так же, как и у бактерий, хотя сама по себе полимераза напоминает соответствующий фермент клеток эукариот.
Большинство археев содержит основной белок деления FtsZ, не не содержит прочих бактериальных или эукариотических белков деления. Более того, Crenarchaeota отличаются от остальных археев тем, что не содержат FtsZ. Это позволяет предполагать, что механизм клеточного деления у археев может сильно отличаться от характерного для всех прочих организмов.
На основании вышесказанного, археи можно рассматривать как некие гибриды со смешанными свойствами, характерными как для эукариот, так и для бактерий, но также обладающие некоторыми уникальными свойствами. Такие свойства археев, вероятно, отражают их положение в системе эволюции. В основном клеточная организация археев напоминает таковую прокариотической клетки, однако многие фундаментальные клеточные события, особенно связанные с реализацией генетической информации, гораздо больше похожи на соответствующие процессы у эукариот, чем у бактерий. В настоящее время исследователи проявляют большой интерес к исследованиям некоторых из этих процессов, поскольку в результате они должны получить много новых сведений о процессе эволюции эукариот. Более того, поскольку эти процессы протекают проще, чем у эукариот, не исключено, что некоторые археи в экспериментальном плане окажутся проще и будут все больше использоваться в качестве модельных систем в исследовании таких процессов, как репликация и транскрипция ДНК.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Сходства и различия эукариот, архей, бактерий
На протяжении многих лет с момента открытия существования микроорганизмов не было однозначного понимания их места в живой природе. Их относили к растениям (отсюда устаревший ныне термин «микрофлора[1]»), затем разделяли на группы среди растений и животных. Сейчас очевидно, что термин «микроорганизм» не имеет систематического смысла, то есть говорит исключительно о микроскопическом размере объекта.
Группы эукариот
В настоящее время микроорганизмы разделяют на две большие группы, принципиально отличающиеся строением клетки – эукариоты и прокариоты (рис. 1). Группа эукариот включает в себя микроскопические водоросли, простейших и микроскопические грибы, такие как дрожжи и плесневые грибы. К прокариотам до 80-х годов относили исключительно бактерий, однако группой исследователей под руководством Карла Вёзе в ходе анализа последовательностей 16S рРНК, было обнаружено, что архебактерии (археи) по своему происхождению являются самостоятельной группой, что подтверждается рядом отличий в их строении и метаболизме: одни черты роднят их с бактериями, другие – с эукариотами, а некоторые являются совершенно уникальными. В частности, первые открытые археи отличаются своей удивительной способностью обитать в экстремальных местах обитания: при высоких температурах, давлении, сильнокислых или сильнощелочшых условиях среды. Например, большинство гипертермофильных архей растут при температуре 80 ℃, а Methanopyrus kandleri – при 122 ℃. Другой пример: рекордсмен среди устойчивых к кислой среде архей растет в условиях, эквивалентных 1,2 М серной кислоте. Для сравнения – содержание соляной кислоты в желудочном соке в норме составляет 0,14 – 0,16 М.
Рисунок 1. Группы микроорганизмов
Сходства и различия в строении клеток прокариот и эукариот
Для существования клеток любого типа, и прокариотических, и эукариотических, необходимо наличие цитоплазматической мембраны, отделяющей клетку от внешней среды; цитоплазмы, заполняющей клетку, а также генетического аппарата и рибосом, позволяющих хранить и реализовывать генетическую информацию. Однако, строение мембраны и рибосом, а также организация генетического материала для этих групп могут различаться (рис.2)
Основное различие прокариот и эукариот состоит в том, что в клетках прокариот генетический материал располагается непосредственно в цитоплазме и представлен нуклеоидом, содержащим чаще всего замкнутую в кольцо молекулу ДНК. У эукариот генетический материал отделен ядерной оболочкой и, соответственно, заключен в ядре. Он представлен линейными молекулами ДНК, «упакованными» в хромосомы.
И у прокариот, и у эукариот есть рибосомы, необходимые для синтеза белка, но рибосомы прокариот меньше эукариотических. Рибосомы бактерий состоят их трех, а не четырех молекул рРНК. Рибосомы архей по некоторым признакам похожи на бактериальные, а по некоторым – на эукариотические. Например, на рибосомы архей не действует антибиотик хлорамфеникол, связывающий рибосомы бактерий, в то время как дифтерийный токсин, останавливающий биосинтез белка у эукариот, действует и на архей.
Кроме рибосом внутри прокариотической клетки нет других органелл и мембранных структур, в то время как эукариотические клетки содержат эндоплазматическую сеть, аппарат Гольджи, митохондрии и другие органеллы. Внутри клеток прокариот могут быть газовые пузырьки или другие включения, окруженные белковой оболочкой.
Рисунок 2. Строение клеток прокариот (на примере бактерий) и эукариот
Такое увеличение площади мембраны необходимо в связи с тем, что энергетические процессы, такие как дыхание и фотосинтез, происходящие у эукариот на внутренних мембранах митохондрий и хлоропластов соответственно, у прокариот происходит непосредственно на мембране клетки.
Цитоскелет прокариот не включает в себя характерных для эукариотической клетки элементов (микротрубочек, актиновых филаментов, микрофиламентов) и образован другими белками. Прокариоты не способны к эндоцитозу и амебоидному движению.
Клеточные покровы прокариот и эукариот так же существенно отличаются. Клетки бактерий и эукариот покрыты цитоплазматической мембраной, состоящей из двойного слоя фосфолипидов, в которых жирные кислоты связаны с молекулой глицерина сложноэфирной связью. Мембранные липиды архей вместо жирных кислот содержат изопреновые цепочки, соединенные с глицерином простой эфирной связью (рис. 3). Липиды такой мембраны зачастую объединяются в один слой с двумя гидрофильными головками и одной гидрофобной «сшивкой» из двух хвостов. Это делает мембрану более устойчивой к экстремальным условиям, в которых обитают некоторые археи.
Рисунок 3. Строение цитоплазматической мембраны бактерий, эукариот и архей
Клеточная стенка бактерий состоит из пептидогликана (муреина), которого нет ни у архей, ни у эукариот. Клетки архей чаще всего покрыты белковым S-слоем, защищающим от воздействия стрессовых условий, а в тех случаях, когда клеточная стенка все-таки присутствует, в ее состав входит похожее по структуре вещество – псевдомуреин.
Отличается и строение жгутиков. Бактериальные жгутики образованы белком флагеллином который, закручиваясь в спираль, формирует полую внутри нить жгутика. Жгутики архей похожи на бактериальные: они приводят клетку в движение, вращаясь по тому же механизму, но они не имеют полости внутри и образованы гликопротеинами. Жгутики эукариот в свою очередь состоят из десяти пар микротрубочек, где одна из пар центральная, а еще девять окружают ее.
Клетки бактерий, архей и эукариот отличаются не только чертами своего строения, существует еще рад биохимических и молекулярных признаков, на которые стоит обратить внимание. Кратко все признаки для каждой группы изложены в таблице 1.
Таблица 1. Сходства и различия в строении клеток бактерий, археи и эукариот
Светочувствительные белки архей и эукариот оказались родственниками
Ученые из Центра исследований молекулярных механизмов старения и возрастных заболеваний МФТИ совместно с коллегами из Германии и США впервые получили и исследовали структуру светоактивируемого протонного насоса из грибов. Ее сравнили с известными структурами белков с такой же функцией. Оказалось, что белок имеет общего предка с протонным насосом микробов без мембран внутри. Однако он отличается от белков бактерий, в клетках которых есть мембраны, но нет ядра. Полученные результаты важны для дальнейших исследований белков в клетках людей. Работа опубликована в журнале Communications Biology.
Живые организмы делятся на три основных группы (домена) по наличию ограниченных мембранами областей в составе клеток. Археи (первый домен) не содержат никаких мембранных частей в клетках. Более сложно устроены бактерии (второй домен) — в их клетках присутствуют органеллы, окруженные мембранами. Эукариоты (третий домен), в свою очередь, содержат и мембранные органеллы, и ядро в составе клеток. К последней группе относятся царства животных (в которое входят и люди), грибов, растений и некоторые одноклеточные организмы. Эволюционные отношения между представителями разных групп живых существ изучаются при помощи исследования и сравнения геномов и белков.
Ранее сравнение геномов многих животных показало, что светочувствительные белки в клетках выполняют чрезвычайно разные функции и присутствуют во всех царствах, а также во многих крупных вирусах. Наиболее распространенное семейство светочувствительных белков — родопсины. Родопсины первого типа состоят из семи пронизывающих мембрану клетки спиралей (Рисунок 1). Белки используют энергию света для активного и пассивного транспорта ионов через мембрану клетки, запуска сигнальных реакций и активации ферментов в клетках. Светочувствительные белки применяются в оптогенетике — методике исследования нервных импульсов, которые можно возбуждать при помощи света. Они считаются распространенными светособирающими белками на Земле и основными «световозахватителями» в океанах. Учитывая повсеместное распространение родопсинов и их важную экологическую роль, нет сомнений в том, что эти белки сыграли значительную роль в эволюции жизни на Земле. На ранних этапах эволюции многие организмы могли использовать родопсины в качестве одного из дополнительных источников энергии, что давало им эволюционное преимущество. Широкое разнообразие протонных помп, а также их древность позволяют исследовать глобальные эволюционные процессы по истории изменений этих белков. Однако родопсины эукариот исследованы менее обширно, так как их выделение и кристаллизация значительно сложнее.
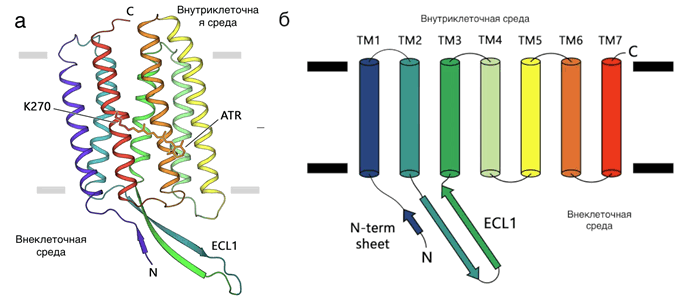
Структура исследованного родопсина и схема соединения спиралей белка. Источник: Communications Biology
Авторы исследования использовали недавно разработанные ими же методы экспрессии (в системе LEXSY) для выделения родопсина 1 типа из одноклеточного гриба. Ученые кристаллизовали и получили структуру белка. Оказалось, что структура родопсина из гриба крайне похожа на структуру протонного насоса археи. Единственное значительное отличие было найдено во внутриклеточной части белка: петля ECL1 гораздо длиннее подобной у родопсина архей (см. рисунок 1). Авторы изучили функции этой петли и обнаружили, что она связывается с внутриклеточным концом, тем самым повышая стабильность белка. Для определения эволюционных отношений между родопсинами ученые сравнили известные структуры и последовательности белков. Оказалось, что последовательности и структуры родопсинов эукариот крайне схожи с таковыми у белков архей. Видимо, гены родопсинов подвергаются обширному горизонтальному переносу между организмами, что усложняет поиск общего предка. Однако высокое структурное сходство протонных помп архей и эукариот, наряду с функциональным сходством, представляет собой убедительное доказательство архейного происхождения эукариотических протонных родопсинов и, скорее всего, всех других эукариотических родопсинов.
«Мы получили первую кристаллическую структуру высокого разрешения светочувствительного протонного насоса из организма гриба и выяснили функциональную роль его N-концевой области. Исследованный родопсин был экспрессирован в системе LEXSY, затем мы его кристаллизовали. Значит, система экспрессии LEXSY может быть сильным инструментом для получения мембранных белков эукариот для структурных исследований. Мы также сравнили последовательность и структуру полученного родопсина со светочувствительными протонными помпами из разных царств. Анализ показал, что эукариотические и архейные родопсины имеют глубокое структурное сходство, что подтверждает гипотезу об архейном происхождении родопсинов, найденных в геноме эукариот. Полученные результаты важны как для понимания эволюции животных, так и для дальнейших исследований родопсинов эукариот», — поясняет Дмитрий Забельский, сотрудник лаборатории химии и физики липидов Центра исследований молекулярных механизмов старения и возрастных заболеваний МФТИ, аспирант Физтех-школы физики и исследований им. Ландау МФТИ.
Работа выполнена при поддержке Министерства науки и высшего образования РФ, а также Российского научного фонда и Российского фонда фундаментальных исследований.
Новооткрытый микроб заполняет брешь между прокариотами и эукариотами
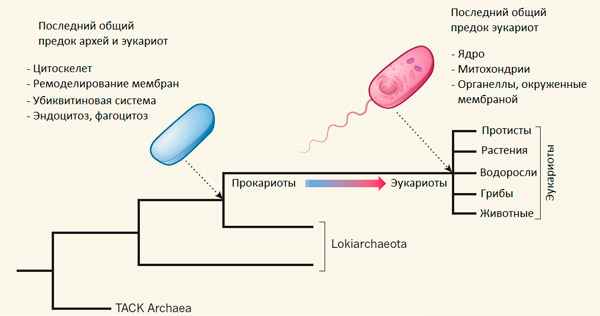
Рис. 1. Положение локиархей (Lokiarchaeota) на эволюционном дереве. Эукариоты — одна из ветвей локиархей. TACK — надтип архей, включающий типы Thaumarchaeota, Aigarchaeota, Crenarchaeota, Korarchaeota. Рисунок из синопсиса к обсуждаемой статье в Nature
В донных осадках на глубине 3283 м в Северном Ледовитом океане обнаружены микроорганизмы из надцарства архей, более близкие к эукариотам, чем любые другие прокариоты. Судя по набору генов, новая группа микробов, получившая название локиархей, обладает многими важными признаками эукариот, включая актиновый цитоскелет и способность к фагоцитозу. Скорее всего, именно благодаря этой способности предкам эукариот удалось захватить бактериального симбионта, давшего начало митохондриям. Открытие подтвердило, что эукариоты не просто имели общего предка с современными археями (что соответствует «трехдоменному» древу жизни), а произошли от одной, вполне определенной, группы архей, что соответствует «двухдоменному» древу и формально (с точки зрения кладистической систематики) заставляет считать эукариот подгруппой архей.
Происхождение эукариот — одна из самых интригующих проблем эволюционной биологии. Для объяснения отдельных этапов становления эукариотической клетки и всего процесса в целом предложены сотни гипотез, как конкурирующих, так и дополняющих друг друга.
На сегодняшний день твердо установлено, что митохондрии и пластиды эукариотической клетки являются потомками симбиотических бактерий (альфапротеобактерий и цианобактерий соответственно). Митохондрии были уже у последнего общего предка всех современных эукариот: это их универсальная черта. Хотя некоторые современные эукариоты лишены митохондрий, это — результат вторичной утраты.
Природа «хозяйской» клетки, некогда захватившей бактериальных симбионтов, менее очевидна, чем происхождение митохондрий и пластид. Геном эукариот явно имеет химерное происхождение: часть генов досталась им от архей, другая — от бактерий (в том числе от симбионтов, но не только от них). Гены архейного происхождения выполняют в эукариотической клетке в основном «центральные» функции (такие как работа с генетической информацией и синтез белка), гены бактериального происхождения — в основном «периферические» (обмен веществ, взаимодействие с внешней средой). По-видимому, предок эукариот (та клетка, которая приобрела митохондриального симбионта) был близок к археям, а бактериальных генов он нахватался путем горизонтального переноса. Недавние исследования показали, что массированное заимствование бактериальных генов происходило на ранних этапах формирования чуть ли не всех крупных клад (групп) архей, так что предок эукариот в этом плане не был исключением (S. Nelson-Sathi et al., 2014. Origins of major archaeal clades correspond to gene acquisitions from bacteria).
Однако эукариотическая клетка устроена намного сложнее, чем прокариотическая, причем далеко не для всех эукариотических генов и молекулярных систем найдены очевидные прокариотические предшественники или аналоги. Откуда взялись эти уникальные особенности эукариотической клетки — вот главный вопрос, на который необходимо ответить. Другой, менее принципиальный, но тоже интересный вопрос, связан со структурой древа жизни (рис. 2). Ставшая уже классической «трехдоменная» система делит все клеточные организмы на три домена (надцарства): бактерии, археи и эукариоты. Альтернативная двухдоменная версия предполагает, что эукариоты отпочковались от одной из групп архей уже после того, как те начали расходиться (дивергировать). В этом случае эукариот формально следует считать подмножеством архей.
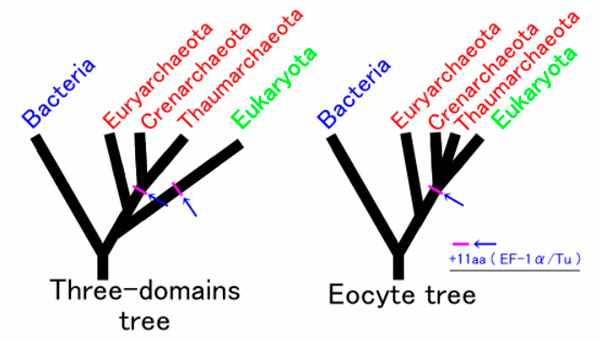
По мере накопления геномных данных позиции двухдоменной модели укрепляются. Становится всё более очевидно, что эукариоты обособились внутри архейной «кроны», то есть являются более близкими родственниками одним археям, чем другим. На роль ближайшего родственника эукариот претендует так называемая клада TACK — надтип архей, включающий типы Thaumarchaeota, Aigarchaeota, Crenarchaeota и Korarchaeota. Впрочем, молекулярно-генетическое сходство эукариот с представителями этой клады недостаточно велико, чтобы однозначно решить вопрос об их родственных связях.
Археи — чрезвычайно разнообразная группа, однако значительная часть их разнообразия сегодня известна ученым лишь по последовательностям гена 16S рРНК, по которому традиционно проводят классификацию прокариот. Это некультивируемые микробы, отказывающиеся расти на лабораторных средах. Из проб, взятых в разных местах (например, из почвы, горячих источников или донных морских отложений) выделяют все имеющиеся там версии гена 16S рРНК и строят по ним эволюционные деревья. На деревьях часто обнаруживаются ветви, не соответствующие ни одной из известных групп прокариот. О такой ветви можно только сказать, что это новая, неизвестная группа, и примерно определить степень ее родства с известными микробами. «Примерно» — потому что одного-единственного гена 16S рРНК недостаточно для более строгих выводов.
В кладе TACK тоже есть такие условные группировки некультивируемых архей, одна из которых называется DSAG (Deep-Sea Archaeal Group — «глубоководная группа архей»). Археи из этой группы обнаружены во многих точках мирового океана на больших глубинах. Фактически, DSAG — одна из самых многочисленных и широко распространенных групп архей в глубоководных донных отложениях, однако ни вырастить чистую культуру этих микробов в лаборатории, ни выделить из проб другие их гены, кроме 16S рРНК, пока не удавалось. Точнее, выделить-то можно, но как понять, какие из бесчисленных фрагментов ДНК в данной пробе принадлежат именно интересующему нас микробу — обладателю необычного варианта 16S рРНК?
Исследователям из Уппсальского университета (Швеция) и Бергенского университета (Норвегия) удалось преодолеть технические препятствия, до недавних пор казавшиеся непреодолимыми, и собрать из кусочков почти полный геном одного микроба из загадочной группы DSAG, а также неполные геномы двух других ее представителей.
Авторы изучили ДНК из пробы грунта, поднятой с глубины 3283 м в Северном Ледовитом океане, недалеко от гидротермальной зоны Loki’s Castle («Замок Локи»), расположенной на склоне срединно-океанического хребта. Анализ последовательностей 16S рРНК показал, что доля архей из группы DSAG в пробе необычайно высока (около 10% всех прокариот и более 70% архей в пробе принадлежат к этой группе). Это и позволило, наряду со сложнейшими современными методами метагеномного анализа и компьютерной обработки геномных данных, отделить фрагменты генетического материала DSAG от всех прочих и собрать из них три генома.
Авторы использовали множество хитроумных приемов. Например, чтобы на первом этапе получить набор фрагментов ДНК, почти наверняка принадлежащих DSAG, они искали «филогенетически значимые» гены (то есть медленно эволюционирующие и при этом имеющиеся почти у всех микробов), строили для каждого гена эволюционное дерево, а затем выбирали те генетические варианты, чье положение на дереве соответствует положению DSAG на дереве 16S рРНК.
Поскольку природа DSAG после сборки геномов во многом прояснилась, авторы присвоили группе более внятное название. Они нарекли глубоководных микробов «локиархеями» (Lokiarchaeota) в честь гидротермального района Замок Локи, а также в связи с тем, что споры мифологов вокруг фигуры этого скандинавского бога сопоставимы по остроте со спорами биологов о происхождении эукариот. Тот представитель группы, чей геном собрали почти полностью, был назван Lokiarchaeum, двух других обозначили пока условно: Loki2 и Loki3.
О чем же рассказали геномы локиархей? Прежде всего, они позволили построить надежное эволюционное дерево, основанное не на единственном гене 16S рРНК, как до сих пор, а на аминокислотных последовательностях 36 консервативных белков, считающихся хорошими «филогенетическими маркерами» (рис. 3).
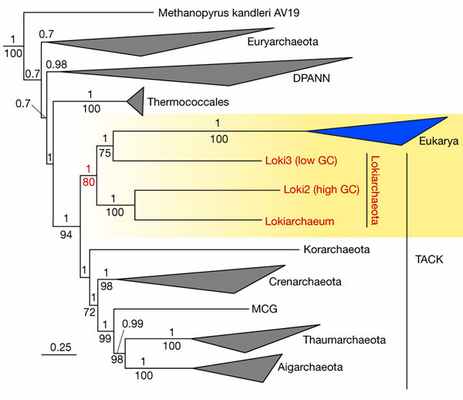
Рис. 3. Эволюционное дерево архей, основанное на последовательностях 36 консервативных белков. Видно, что локиархеи (Lokiarchaeota) образуют единую группу в пределах надтипа TACK, в состав которой входят также эукариоты (Eukarya). Числа над и под ветвями дерева соответствуют двум количественным оценкам достоверности данной ветви. Верхнее число — байесовская апостериорная вероятность (Bayesian posterior probability, см. Bayesian inference), показаны значения выше 0,7; нижнее число — максимально правдоподобное (см. Метод максимального правдоподобия) опорное значение бутстрэпа (bootstrap support value, см. Bootstrapping), показаны значения выше 70. Рисунок из обсуждаемой статьи в Nature
На этом дереве локиархеи образуют единую монофилетическую группу в пределах надтипа TACK. Когда авторы включили в анализируемый массив данные по эукариотам, те пристроились к локиархеям в качестве одной из веточек: ближе к Loki3, чем к Lokiarchaeum и Loki2. Таким образом, локиархеи оказались ближе к эукариотам, чем к другим археям (подобно тому, как шимпанзе ближе к людям, чем к другим обезьянам). С точки зрения кладистической систематики это означает, что эукариоты — подмножество архей (подобно тому, как люди — подмножество обезьян). Таким образом, подтвердилась двухдоменная, а не трехдоменная структура древа жизни.
Одной из особенностей геномов локиархей является повышенная доля генов бактериального происхождения. Это согласуется со сказанным выше о периодическом массовом заимствовании бактериальных генов археями, в том числе, возможно, и предками эукариот.
Если локиархеи — ближайшая родня эукариот, то у них могут найтись гены и признаки, характерные для эукариот, но отсутствующие у других прокариот. И действительно, в геноме Lokiarchaeum нашлось целых 175 белок-кодирующих генов (3,3% от общего числа генов в геноме), похожих на эукариотические, в том числе на так называемые ESPs (eukaryotic signature proteins) — белки, считающиеся уникальными для эукариот. Этот факт, наряду с вышеупомянутым эволюционным деревом, — второй весомый довод в пользу близкого родства локиархей и эукариот.
Авторы скрупулезно разбирают факты, позволяющие утверждать, что «эукариотические» гены Lokiarchaeum не являются результатом случайного загрязнения собранного генома нуклеотидными последовательностями эукариот. Во-первых, эти гены у Lokiarchaeum всегда соседствуют с типично прокариотическими генами, а во многих случаях они находились на прочтенных фрагментах ДНК вместе с генами, уникальными для архей. Во-вторых, эти гены присутствуют в пробе во множестве экземпляров. При этом настоящая эукариотическая ДНК там обнаружена лишь в следовых количествах, а характерные для эукариот гены 18S рРНК вовсе не найдены. В третьих, на эволюционных деревьях «эукариотические» гены локиархей во многих случаях образуют базальные ветви, то есть отделяются от общего ствола до начала дивергенции эукариот. Это значит, что они унаследованы от общего с эукариотами предка, а не заимствованы у эукариот позднее.
Среди «эукариотических» генов локиархей наибольший интерес представляют гены, связанные с подвижностью клетки и ее мембраны, с возможностью формирования разнообразных мембранных структур и активного захвата объектов из внешней среды (см. фагоцитоз). Ключевую роль в выполнении этих функций у эукариот играет белок актин — важнейший компонент цитоскелета. В геноме Lokiarchaeum имеется целых пять генов, кодирующих белки, похожие на эукариотические актины и актиноподобные белки (ARPs, actin-related proteins). Эти белки локиархей («локиактины») намного ближе к актинам эукариот, чем открытые ранее у других архей гомологи актина — так называемые кренактины. Эволюционные деревья показывают, что общий предок эукариот уже имел более одного актинового гена, то есть начало диверсификации актинов предшествовало появлению эукариот. Кроме того, у локиархей есть белки, похожие на известные эукариотические регуляторы формирования актиновых нитей. Таких белков нет у других прокариот. По мнению авторов, эти факты показывают, что у локиархей с большой вероятностью имеется актиновый цитоскелет.
У локиархей также обнаружено большое разнообразие особых регуляторных белков (малых ГТФаз из надсемейства Ras, см. Ras superfamily), играющих у эукариот важную роль в регуляции работы актинового цитоскелета в ходе таких процессов, как фагоцитоз и везикулярный транспорт. У некоторых других прокариот тоже найдены похожие белки, но в несопоставимо меньшем количестве.
Еще одна «эукариотическая» черта локиархей — присутствие особого комплекса генов (ESCRT), служащего для образования всевозможных изгибов клеточной мембраны и отпочковывания мембранных пузырьков. У эукариот комплекс ESCRT обслуживает, помимо прочего, систему убиквитин-опосредованной деградации белка (см. Убиквитин); в геномах локиархей обнаружены компоненты и этой системы тоже.
Рибосомы локиархей, по-видимому, больше похожи на рибосомы эукариот, чем любые другие прокариотические рибосомы. В частности, только у локиархей есть «эукариотический» рибосомный белок L22e.
Таким образом, локиархеи оказались своеобразными переходными формами, заполняющими брешь между типичными про- и эукариотами. Важность этого открытия для решения проблемы происхождения эукариот трудно переоценить. Анализ генома локиархей и его сравнение с геномами других представителей клады TACK показали, что архейный предок эукариот имел сложное строение и обладал многими продвинутыми признаками, которые раньше считались уникальными для эукариот. Такие эукариотические «ноу-хау», как убиквитиновая система деградации отслуживших белков, актиновый цитоскелет, везикулярный транспорт, а возможно, и фагоцитоз, необходимый для захвата симбионтов, — всё это, вероятно, уже имелось у тех архей, которые дали начало эукариотам, подружившись с альфапротеобактерией — предком будущих митохондрий.
В свете новых данных становится понятнее отмеченное ранее мозаичное распределение отдельных эукариотических признаков у архей из клады TACK. Скорее всего, эти признаки имелись у их общего предка с локиархеями и эукариотами, но потом в разных ветвях они многократно и независимо терялись (рис. 4).

Рис. 4. Распределение эукариотических признаков в разных группах архей. Дерево слева показывает предполагаемую последовательность приобретения признаков (цветные кружки — приобретение признака, перечеркнутые кружки — утрата). Наполовину закрашенные кружки означают, что данный признак есть лишь у некоторых представителей группы. Рисунок из обсуждаемой статьи в Nature
Растущие возможности метагеномики и компьютерного анализа нуклеотидных последовательностей позволяют надеяться, что среди некультивируемых микробов — этой «темной материи» мира прокариот — будет найдено еще немало интересных форм. В частности, среди локиархей (которые, как мы знаем, являются широко распространенной и массовой глубоководной группой) вполне могут обнаружиться виды, стоящие еще ближе к эукариотам, чем описанные в обсуждаемой статье.
Источники:
1) Anja Spang, Jimmy H. Saw, Steffen L. Jørgensen, Katarzyna Zaremba-Niedzwiedzka, Joran Martijn, Anders E. Lind, Roel van Eijk, Christa Schleper, Lionel Guy & Thijs J. G. Ettema. Complex archaea that bridge the gap between prokaryotes and eukaryotes // Nature. Published online 06 May 2015.
2) T. Martin Embley & Tom A. Williams. Steps on the road to eukaryotes // Nature. Published online 06 May 2015.
Читайте также: