Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Добавил пользователь Владимир З. Обновлено: 29.01.2026
Различия в концентрации ионов на противоположных сторонах клеточной мембраны приводят к напряжению, названному мембранным потенциалом. Много ионов имеют градиент концентрации поперек мембраны, включая калий (K+), который является в высокой внутренней части и низкой концентрации вне мембраны. Натрий (Na+) и хлорид (Замкнутый–) ионы - при высоких концентрациях во внеклеточной области, и низких концентрациях во внутриклеточных областях. Эти градиенты концентрации обеспечивают потенциальную энергию вести формирование мембранного потенциала. Это напряжение установлено, когда мембрана имеет проходимость к одному или более ионам. В самом простом случае, иллюстрированном здесь, если мембрана выборочно водопроницаема к калию, они положительно обвиняли, что ионы могут распространить вниз градиент концентрации к внешней стороне ячейки, оставляя позади неданные компенсацию отрицательные обвинения. Это разделение обвинений-, каков вызывает мембранный потенциал. Отметьте, что оптовые решения любой стороны мембраны electo-нейтральны. Аналогично, система в целом нейтральна гальваностереотипом. "Неданные компенсацию" положительные обвинения вне ячейки, и неданных компенсацию отрицательных обвинений в ячейке, физически выстраиваются в линию на мембранной поверхности и привлекают друг друга поперек мембраны. Таким образом, мембранный потенциал физически расположен только в непосредственной близости мембраны. Это - разделение этих обвинений поперек них мембрана, которая является основанием мембранного напряжения. Отметьте также, что эта диаграмма - только приближение ионных вкладов в мембранный потенциал. Другие ионы, включая натрий, хлорид, кальций и другие играют более незначительную роль, даже при том, что они имеют сильные градиенты концентрации, потому что они более ограничили проходимость чем калий. Ключ: Синие пятиугольники - ионы натрия; Фиолетовые площади - ионы калия; Желтые круги - ионы Choloride; Оранжевые прямоугольники - Анионы (они являются результатом разнообразия источников, включая белки). Большая фиолетовая структура со стрелкой представляет трансмембранный канал калия и руководство чистого движения калия.
Роль мембраны в первую очередь состоит в создании препятствия к смешиванию растворов, расположенных по её разные стороны. Мембрана может быть либо электрически индифферентной, диффузия через которую возможна для всех частиц, имеющихся в растворе; либо полупроницаемой (активной), через такую мембрану некоторые частицы пройти не могут (см.
Содержание
Введение
Мембранный потенциал (также трансмембранное потенциальное или мембранное напряжение) - различие в электрическом потенциале между интерьером и внешностью биологической клетки. Все клетки животных окружены плазменной мембраной, составленной из двойного слоя липида с разнообразием типов белков, вложенных в это. Мембранный потенциал возникает прежде всего из взаимодействия между мембраной и действиями двух типов трансмембранных белков, вложенных в плазменную мембрану. Мембрана служит и изолятором и барьером распространения к движению ионов. Белки транспортера/насоса иона активно выдвигают ионы поперек мембраны, чтобы устанавливать градиенты концентрации поперек мембраны, и каналы иона позволяют ионам перемещать поперек мембраны вниз те составляющие концентрации. Этот процес известен как облегченное распространение. В самом фундаментальном примере этого, транспортер иона Na+/K+-ATPase качает ионы натрия от внутренней части до внешней стороны, и ионов калия от внешней стороны до внутренней части клетки. Это устанавливает два градиента концентрации: градиент для натрия, где его концентрация намного выше снаружи чем в ячейке, и градиенте для калия, где его концентрация намного выше в ячейке чем снаружи.
Трансмембранные отборные калием каналы утечки позволяют ионам калия распространяться поперек мембраны, вниз градиента концентрации, который был установлен ATPase, создавая разделение обвинения, и таким образом, напряжение, поперек мембраны.
Трансмембранные отборные калием каналы утечки позволяют ионам калия распространяться поперек мембраны, вниз градиента концентрации, который был установлен ATPase, создавая разделение обвинения, и таким образом, напряжение, поперек мембраны. В почти во всех случаях, ион, который определяет так называемый "отдыхающий" мембранный потенциал ячейки, является калием, хотя другие ионы действительно вносят вклад большим количеством незначительных способов. В соответствии с соглашением, признак мембранного потенциала определяется как напряжение внутри относительно основания вне клетки. В случае калия, его распространение вниз его градиента концентрации, к внешней стороне ячейки, создает трансмембранное напряжение, которое является отрицательным относительно внешней стороны ячейки, и типично–60 к–80 милливольтам (милливольт) в амплитуде.
Фактически все eukaryotic клетки (включая клетки от животных, заводов, и грибов) поддерживают трансмембранный потенциал отличный от нуля, обычно с отрицательным напряжением в интерьере клетки по сравнению с внешностью клетки. Мембранный потенциал имеет две основных функции. Сначала он позволяет клетке функционировать как батарея, обеспечивая власть управления разнообразием "молекулярных устройств" вложенных в мембрану. Во вторых, в электрически легковозбудимых клетках, типа нейронов и клеток мускула, это используется для того, чтобы передать сигналы между различными частями клетки. Сигналы произведены, открывая или закрывая каналы иона однажды в мембране, производя местное изменение в мембранном потенциале, который заставляет электрический ток течь быстро к другим пунктам в мембране.
В нелегковозбудимых клетках, и в легковозбудимых клетках в их местах основания, мембранный потенциал проведен по относительно устойчивой ценности, названной отдыхающим потенциалом. Для нейронов, типичные ценности отдыхающего потенциального диапазона от–70 до–80 милливольт; то есть, интерьер клетки имеет отрицательное напряжение основания немного меньше, чем одна десятая вт. Открытие и закрытие каналов иона могут вызвать отъезд от отдыхающего потенциала. Это называют деполяризацией, если внутреннее напряжение становится более уверенным (скажем от-70 милливольтов до-60 милливольтов), или гиперполяризация, если внутреннее напряжение становится более отрицательным (говорят от-70 милливольтов до-80 милливольтов). В легковозбудимых клетках, достаточно большая деполяризация может вызвать потенциал действия, в котором мембранный потенциал изменяется быстро и значительно в течение короткого времени (на заказе в пределах от 1 - 100 миллисекунд), часто полностью изменяя его полярность. Потенциалы действия произведены активацией определенных каналов иона напряжения-gated.
В нейронах, факторы, которые влияют на мембранный потенциал, разнообразны. Они включают многочисленные типы каналов иона, некоторые, которые являются химически gated и некоторые, которые являются напряжением-gated. Поскольку каналами иона напряжения-gated управляет мембранный потенциал, в то время как мембранный сам потенциал - под влиянием этих тех же самых каналов иона, петли обратной связи, которые учитывают сложную временную динамику, возникают дополнительно
Потенциал покоя и потенциал действия
Мембрана всех живых клеток поляризована. Внутренняя сторона мембраны несет отрицательный заряд по сравнению с межклеточным пространством (рис. 1). Величина заряда, который несет мембрана называется мембранным потенциалом (МП). В невозбудимых тканях МП низкий, и составляет около -40 мВ. В возбудимых тканях он высокий, около -60 - -100 мВ и называется потенциалом покоя (ПП).
Потенциал покоя, как и любой мембранный потенциал формируется за счет избирательной проницаемости клеточной мембраны. Как известно, плазмолемма состоит из липидного бислоя, через который движение заряженных молекул затруднено. Белки, встроенные в мембрану, могут избирательно изменять проницаемость мембраны для различных ионов, в зависимости от приходящих стимулов. При этом, для формирования потенциала покоя ведущую роль играют ионы калия, кроме них важны ионы натрия и хлора.
Рис. 1. Концентрации и распределение ионов с внутренней и внешней стороны мембраны.
Большинство ионов распределяются неравномерно с внутренней и внешней стороны клетки (рис. 1). Внутри клетки концентрация ионов калия выше, а натрия и хлора – ниже, чем снаружи. В состоянии покоя мембрана проницаема для ионов калия и практически непроницаема для ионов натрия и хлора. Несмотря на то, что калий может свободно выходить из клетки, его концентрации остаются неизменными благодаря отрицательному заряду на внутренней стороне мембраны. Таким образом, на калий действуют две силы, находящиеся в равновесии: осмотические (градиент концентрации К + ) и электрические (заряд мембраны), благодаря чему число входящих в клетку ионов калия равно выходящим. Движение калия осуществляется через калиевые каналы утечки, открытые в состоянии покоя. Величину заряда мембраны, при которой ионы калия находятся в равновесии можно вычислить по уравнению Нернста:
где Ек — равновесный потенциал для К + ; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; n — валентность К + (+1), [К + н] — [К + вн] — наружная и внутренняя концентрации К + .
Если подставить в уравнение значения из таблицы на рис. 43, то мы получим величину равновесного потенциала, равную примерно -95 мВ. Это значение вписывается в диапазон мембранного потенциала возбудимых клеток. Отличия ПП разных клеток (даже возбудимых) могут возникать по трем причинам:
- отличия внутриклеточной и внеклеточной концентраций ионов калия в разных тканях (в таблице приведены данные по среднестатистическому нейрону);
- натрий-калиевая АТФаза может вносить свой вклад в значение заряда, так как она выводит из клетки 3 Na + в обмен на 2 К + ;
- несмотря на минимальную проницаемость мембраны для натрия и хлора, эти ионы все-таки могут попадать в клетки, хоть и от 10 до 100 раз хуже, по сравнению с калием.
Чтобы учесть проникновение других ионов в клетку существует уравнение Нернста-Гольдмана:
, где Еm — мембранный потенциал; R — газовая постоянная; Т — абсолютная температура; F — число Фарадея; РK , PNa и РCl — константы проницаемости мембраны для К + Na + и Сl, соответственно; [К + н], [K + вн], [Na + н], [Na + вн], [Сl — н] и [Сl — вн ]- концентрации K + , Na + и Сl снаружи (н) и внутри (вн) клетки.
Такое уравнение позволяет установить более точную величину ПП. Обычно, мембрана оказывается на несколько мВ менее поляризована, по сравнению с равновесным потенциалом для К + .
Потенциал действия (ПД) может возникать в возбудимых клетках. Если на нерв или мышцу нанести раздражение выше порога возбуждения, то ПП нерва или мышцы быстро уменьшится и на короткий промежуток времени (миллисекунда) произойдет кратковременная перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной, после чего восстановится ПП. Это кратковременное изменение ПП, происходящее при возбуждении клетки называется потенциалом действия.
Возникновение ПД возможно благодаря тому, что в отличие от ионов калия, ионы натрия далеки от равновесия. Если подставить в уравнение Нернста натрий вместо калия, то мы получим равновесный потенциал, равный примерно +60 мВ. Во время ПД, происходит кратковременное увеличение проницаемости для Na + . При этом, натрий начнет проникать в клетку под действием двух сил: по градиенту концентрации и по заряду мембраны, стремясь подстроить заряд мембраны под свой равновесный потенциал. Движение натрия осуществляется по потенциал-зависимым натриевым каналам, которые открываются в ответ на смещение мембранного потенциала, после чего сами инактивируются.
Рис. 2. Потенциал действия нервного волокна (А) и изменение проводимости мембраны для ионов натрия и калия (Б).
На записи ПД выглядит как кратковременный пик (рис. 44), имеющий несколько фаз.
- Деполяризация (фаза нарастания) (рис. 44) – увеличение проницаемости для натрия из-за открытия натриевых каналов. Натрий стремится к своему равновесному потенциалу, но не достигает его, так как канал успевает инактивироваться.
- Реполяризация – возвращение заряда к величине потенциала покоя. Помимо калиевых каналов утечки здесь подключаются потенциал-зависимые калиевые каналы (активируются от деполяризации). В это время калий выходит из клетки, возвращаясь к своему равновесному потенциалу.
- Гиперполяризация (не всегда) – возникает в случаях, если равновесный потенциал по калию превышает по модулю ПП. Возвращение к ПП происходит после возвращения к равновесному потенциалу по К + .
Во время ПД происходит изменение полярности заряда мембраны. Фаза ПД, при которой заряд мембраны положителен, называется овершутом (рис. 2).
Благодаря Н-воротам инактивация канала происходит раньше, чем потенциал на мембране достигнет равновесной величины по натрию. После прекращения поступления натрия в клетку, происходит реполяризация за счет выходящих из клетки ионов калия. При этом к каналам утечки в этом случае подключаются еще и потениал-активируемые калиевые каналы. Во время реполяризации, в быстром натриевом канале быстро закрываются М-ворота. Н-ворота открываются гораздо медленнее и остаются закрытыми еще некоторое время после возвращения заряда к потенциалу покоя. Этот период принято называть периодом рефрактерности.
Рис. 3. Работа потенциал-управляемого натриевого канала.
Концентрации ионов внутри клетки восстанавливает натрий-калиевая АТФаза, которая с затратой энергии в виде АТФ откачивает из клетки 3 иона натрия и закачивает 2 иона калия.
По немиелинизированному волокну или по мембране мышцы потенциал действия распространяется непрерывно. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до порогового значения, в результате чего на соседнем участке возникает деполяризация. Главную роль в возникновении потенциала на новом участке мембраны предыдущий участок. При этом на каждом участки сразу после ПД наступает период рефрактерности, за счет которое ПД распространяется однонаправленно. При прочих равных условиях распространение потенциала действия по немиелинизированному аксону происходит тем быстрее, чем больше диаметр волокна. У млекопитающих скорость составляет 1-4 м/с. Поскольку у беспозвоночных животных отсутствует миелин, в гигантских аксонах кальмара скорость ПД может достигать 100 м/c.
По миелинизированному волокну потенциал действия распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах немиелинизированных волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до порогового значения, что приводит к возникновению в них новых потенциалов действия, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2-й, 3-й, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Сальтаторное проведение увеличивает скорость проведения ПД 15-20 раз до 120 м/с.
Работа нейронов
Нервная система состоит из нейронов и глиальных клеток. Однако, главную роль в проведении и передаче нервных импульсов играют нейроны. Они получают информацию от множества клеток по дендритам, анализируют ее и передают или не передают на следующий нейрон.
Передача нервного импульса с одной клетки на другую осуществляется с помощью синапсов. Различают два основных типа синапсов: электрические и химические (рис. 4). Задача любого синапса – передать информацию с пресинаптической мембраны (мембрана аксона) на постсинаптическую (мембрана дендрита, другого аксона, мышцы или другого органа-мишени). Большинство синапсов нервной системы образуется между окончанием аксонов и дендритами, которые в области синапса образуют дендритные шипики.
Преимущество электрического синапса состоит в том, что сигнал с одной клетки на другую переходит без задержки. Кроме того, такие синапсы не утомляются. Для этого пре- и постсинаптические мембраны соединены поперечными мостиками, через которые ионы из одной клетки могут перемещаться в другую. Однако, существенным минусом такой системы является отсутствие однонаправленной передачи ПД. То есть, он может передаваться как с пресинаптической мембраны на постсинаптическую, так и наоборот. Поэтому, такая конструкция встречается достаточно редко и в основном – в нервной системе беспозвоночных.
Рис. 4. Схема строения химического и электрического синапсов.
Химический синапс весьма распространен в природе. О устроен сложнее, так как необходима система преобразования электрического импульса в химический сигнал, затем, вновь в электрический импульс. Все это приводит к возникновению синаптической задержки, которая может составить 0,2-0,4 мс. Кроме того, может произойти истощение запасов химического вещества, что приведет к утомлению синапса. Однако, такой синапс обеспечивает однонаправленность передачи ПД, что является его главным преимуществом.
Рис. 5. Схема работы (а) и электронная микрофотография (б) химического синапса.
В состоянии покоя окончание аксона, или пресинаптическое окончание, содержит мембранные пузырьки (везикулы) с нейромедиатором. Поверхность везикул заряжена отрицательно, чтобы предотвратить связывание с мембраной, и покрыта специальными белками, и принимающими участие в высвобождении везикул. В каждом пузырьке находится одинаковое количество химического вещества, которое называется квантом нейромедиатора. Нейромедиаторы весьма разнообразны по химическому строению, однако, большинство из них производятся прямо в окончании. Поэтому, в нем могут находиться системы, для синтеза химического посредника, а также аппарат Гольджи и митохондрии.
Постсинаптическая мембрана содержит рецепторы к нейромедиатору. Рецепторы могут быть в виде как ионных каналов, открывающихся при контакте со своим лигандом (ионотропные), так и мембранными белками, запускающими внутриклеточный каскад реакций (метаботропные). Один нейромедиатор может иметь несколько как ионотропных, так и метаботропных рецепторов. При этом, часть из них может быть возбуждающими, а часть – тормозными. Таким образом, реакцию клетки на нейромедиатор будет определять тип рецептора на ее мембране, и разные клетки могут совершенно по-разному реагировать на одно и то же химическое вещество.
Между пре- и постсинаптической мембраной располагается синаптическая щель, шириной 10-15 нм.
При приходе ПД на пресинаптическое окончание, на нем открываются потенциал-активируемые кальциевые каналы и ионы кальция входят в клетку. Кальций связывается с белками на поверхности везикул, что приводит к их транспортировке к пресинаптической мембране с последующим слиянием мембран. После такого взаимодействия нейромедиатор оказывается в синаптической щели (рис. 5) и может связаться со своим рецептором.
Ионотропные рецепторы – это лиганд-активируемые ионные каналы. Это значит, что канал открывается только в присутствии определенного химического вещества. Для разных нейромедиаторов это могут быть натриевые, кальциевые или хлорные каналы. Ток натрия и кальция вызывает деполяризацию мембраны, поэтому такие рецепторы называют возбуждающими. Хлорный ток приводит к гиперполяризации, что затрудняет генерацию ПД. Следовательно, такие рецепторы называют тормозными.
Метаботропные рецепторы к нейромедиаторам относят к классу рецепторов, ассоцированных с G-белками (GPCR). Эти белки запускают разнообразные внутриклеточные каскады реакций, приводящих в конечном итоге либо к дальнейшей передачи возбуждения, либо к торможению.
После передачи сигнала необходимо быстро удалить нейромедиатор из синаптической щели. Для этого в щели присутствуют либо ферменты расщепляющие, нейромедиатор, либо на пресинаптическом окончании или соседних глиальных клетках могут располагаться транспортеры, закачивающие медиатор в клетки. В последнем случае он может использоваться повторно.
Каждый нейрон получает импульсы от 100 до 100 000 синапсов. Одиночная деполяризация на одном дендрите не приведет к дальнейшей передаче сигнала. На нейрон могут приходит одновременно множество как возбуждающих, так и тормозных стимулов. Все они суммируются на соме нейрона. Такая суммация называется пространственной. Далее, может возникнуть или не возникнуть (в зависимости от пришедших сигналов) ПД в области аксонного холмика. Аксонный холмик – это область аксона, примыкающая к соме и обладающая минимальным порогом ПД. Далее импульс распространяется по аксону, конец которого может сильно ветвиться и образовывать синапсы со множеством клеток. Помимо пространственной, существует временная суммация. Она происходит в случае, поступления часто повторяющихся импульсов от одного дендрита.
Помимо классических синапсов между аксонами и дендритами или их шипиками, существуют также синапсы, модулирующие передачу в других синапсах (рис. 6). К ним относят аксо-аксональные синапсы. Такие синапсы способны усиливать или тормозить синаптическую передачу. То есть, если на окончание аксона, образующего аксо-шипиковый синапс, пришел ПД, а в это время по аксо-аксональному синапсу на него пришел тормозный сигнал, высвобождения нейромедиатора в аксо-шипиковом синапсе не произойдет. Аксо-дендритные синапсы могут изменять проведение мембраной ПД на пути от шипика к соме клетки. Также существуют аксо-соматические синапсы, которые могут влиять на суммацию сигнала в области сомы нейрона.
Таким образом, существует огромное многообразие различных синапсов, отличающихся по составу нейромедиаторов, рецепторов и их местоположению. Все это обеспечивает разнообразие реакций и пластичность нервной системы.
Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Ткани и органы. Нервная ткань
Потенциал покоя и потенциал действия
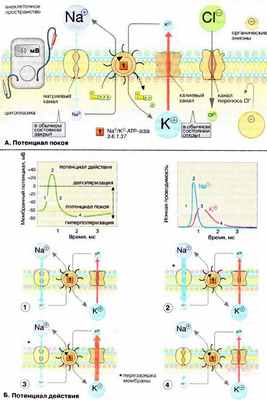
А. Потенциал покоя
Мембраны, в том чикле плазматические, в принципе непроницаемы для заряженных частиц. Правда, в мембране имеется Na + /K + -АТФ-аза (Nа + /К + -АТР-аза), осуществляющая активный перенос ионов Na + из клетки в обмен на ионы К + . Этот транспорт энергозависим и сопряжен с гидролизом АТФ (АТР) (см. рис. 221). За счет работы «Nа + ,К + -насоса» поддерживается неравновесное распределение ионов Na + и К + между клеткой и окружающей средой (см. с. 128). Поскольку расщепление одной молекулы АТФ обеспечивает перенос трех ионов Na + (из клетки) и двух ионов К + (в клетку), этот транспорт электрогенен, т. е. цитоплазма клетки заряжена отрицательно по отношению к внеклеточному пространству.
Электрохимический потенциал. Содержимое клетки заряжено отрицательно по отношению к внеклеточному пространству. Основная причина возникновения на мембране электрического потенциала (мембранного потенциала Δψ, см. c. 128) — существование специфических ионных каналов. Транспорт ионов через каналы происходит по градиенту концентрации или под действием мембранного потенциала. В невозбужденной клетке часть К + -каналов находится в открытом состоянии и ионы К + постоянно диффундируют из нейрона в окружающую среду (по градиенту концентрации). Покидая клетку, ионы К + уносят положительный заряд, что создает потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости различных ионов (см. таблицу на рис. 129) видно, что каналы, проницаемые для Na + и Cl - , преимущественно закрыты. Ионы фосфата и органические анионы, например белки, практически не могут проходить через мембраны. С помощью уравнения Нернста (см. рис. 129) можно показать, что мембранный потенциал нервной клетки в первую очередь определяется ионами К + , которые вносят основной вклад в проводимость мембраны.
Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na + , К + , Са 2+ и Cl - . Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (или электровозбудимые), например быстрые Na + -каналы, и лиганд-управляемые (или хемовозбудимые), например никотиновые холинэргические рецепторы. Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами (см. рис. 343), белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — «закрыт» — и т.д.).
Б. Потенциал действия
Возбуждение нервной клетки под действием химического сигнала (реже электрического импульса) приводит к возникновению потенциала действия . Это означает, что потенциал покоя -60 мВ скачком изменяется на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с открывания Nа + -канала ( 1 ). Ионы Na + устремляются в клетку (по градиенту концентрации), что вызывает локальное обращение знака мембранного потенциала ( 2 ). При этом Na + -каналы тотчас закрываются, т. е. поток ионов Na + в клетку длится очень короткое время ( 3 ). В связи с изменением мембранного потенциала открываются (на несколько мс) потенциал-управляемые К + -каналы ( 2 ) и ионы К + устремляются в обратном направлении, из клетки. В результате мембранный потенциал принимает первоначальное значение ( 3 ), и даже превышает на короткое время потенциал покоя ( 4 ). После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану проходит небольшая часть ионов Na + и К + , и концентрационные градиенты обоих ионов сохраняются (в клетке выше уровень К + , а вне клетки выше уровень Na + ). Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно. Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.
Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Плазматическая мембрана всех клеток, в том числе и нейронов, представлена двойным слоем (бислоем) фосфолипидов, в котором «головки» фосфолипидов обращены к водной среде внеклеточных и внутриклеточных пространств, а парные «хвосты» погружены внутрь и образуют липидную мембрану. Фосфатный слой растворим в воде (гидрофильный, или полярный), а двойной липидный слой — нерастворим (гидрофобный, или неполярный).
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl - ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl - ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl - на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
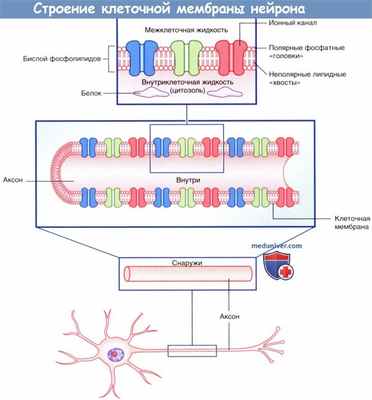
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl - . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl - намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
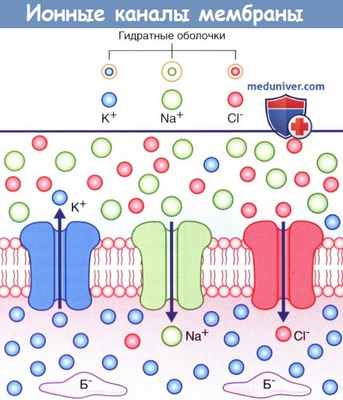
В состоянии покоя ионы Na + и Cl - сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б - ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl - во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl - — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
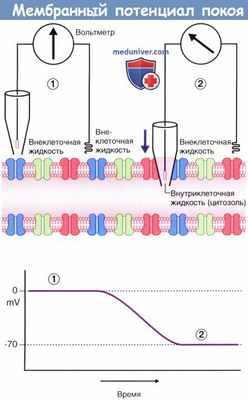
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl - :
Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl - внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
Уравнение Гольдмана сходно с уравнением Нернста, однако в нем перемножаются проницаемость мембраны для каждого иона и его концентрация; это уравнение применяют для установления потенциала «реверсии» через плазматическую мембрану с учетом ее проницаемости для разных ионов. Влияние ионов С1" на потенциал покоя имеет второстепенное значение, поскольку равновесный потенциал и потенциал покоя ионов Cl - приблизительно равны. Значение потенциала покоя в среднем составляет -70 мВ, и обеспечивается за счет суммарного влияния ионов К + и Na + .
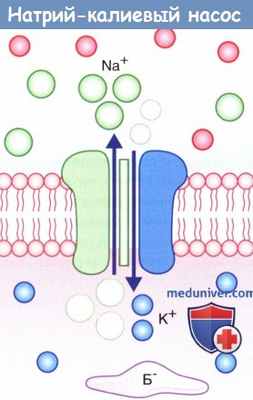
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б - — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.
ОСНОВНЫЕ СВОЙСТВА НЕРВНОЙ КЛЕТКИ
Все живые клетки обладают способностью под влиянием раздражителей переходить из состояния физиологического покоя в состояние активности, или возбуждения. Возбуждение — это комплекс активных электрических, химических и функциональных изменений в нервной, мышечной или железистой ткани, которыми ткань отвечает на внешнее воздействие. Особо важную роль при возбуждении играют электрические процессы, обеспечивающие проведение возбуждения по волокнам и приводящие ткани в рабочее (активное) состояние. Основные закономерности возникновения и протекания процессов возбуждения будут рассмотрены на примере работы нейрона.
Как понять, что электрод попал в клетку? Для того чтобы это понять, нужно знать очень важное свойство всех клеточных структур. Это свойство заключается в том, что внутренняя поверхность клетки всегда заряжена отрицательно по отношению к внешней ее стороне. Поэтому между внешней поверхностью клетки, заряженной электроположительно по отношению к протоплазме, и внутренней стороной клеточной мембраны существует разность потенциалов, которая колеблется в пределах от 60 до 70 мВ. (По данным П. Г. Костюка, у нервной клетки эта разность колеблется в пределах от 30 до 70 мВ.) Разность потенциалов между внешней и внутренней сторонами мембраны клетки, находящейся в невозбужденном состоянии, принято называть мембранным потенциалом, или потенциалом покоя. Как только электрод прокалывает покрывающую клетку мембрану, луч осциллографа сразу отклоняется вниз от своего исходного положения и устанавливается на новом уровне, показывая этим существование разности потенциалов между поверхностью и содержимым клетки (рис. 4).
Мембранный потенциал покоя присутствует на мембране до тех пор, пока клетка жива, и исчезает только с ее гибелью. О существовании этой разности потенциалов было известно давно, только обнаруживали ее иным способом. Л. Гальвани еще в 1794 году показал, что если повредить нерв или мышцу, сделав поперечное сечение и приложив к поврежденной части и к месту повреждения электроды, соединенные с гальванометром, то гальванометр покажет ток, который всегда течет от неповрежденной части ткани к месту разреза. Это течение назвали током покоя.
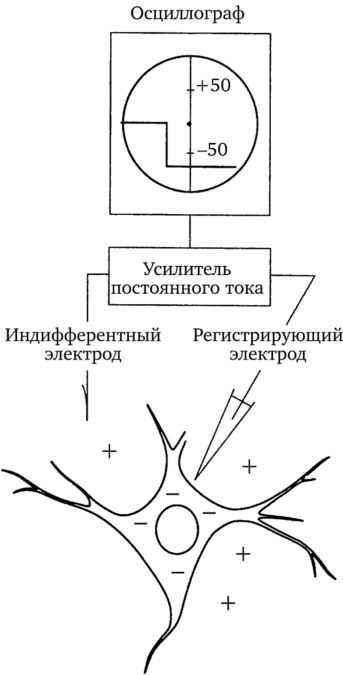
Рис. 4. Мембранный потенциал (потенциал покоя клетки)
Как только микроэлектрод проник в тело клетки, луч осциллографа отклонился до отметки — 60 мВ. Это и есть потенциал покоя клетки, заряженной отрицательно по отношению к «положительной» внеклеточной среде. Разность потенциалов между внутренней и внешней поверхностями клеточной мембраны будет существовать до тех пор, пока клетка жива, потому что электрическая поляризация мембраны — это главное условие, обеспечивающее возбудимость клетки.
По своей физиологической сути ток покоя и мембранный потенциал покоя — это одно и то же. Просто в данном случае электрод не прокалывает ткань, а регистрирует электрический заряд через место ее повреждения. Однако, измеряя разность потенциалов таким способом, физиолог получает несколько меньшие значения — что-то около 30— 50 мВ, поскольку при повреждении часть тока не пойдет через измерительный прибор, а шунтируется (закоротится) в межклеточном пространстве и окружающей исследуемую структуру жидкости. Разность потенциалов можно рассчитать по формуле Нернста:
где R — газовая постоянная; F — число Фарадея; Т — абсолютная температура; [К + ]вн и [К + ]нар — концентрация калия соответственно внутри и снаружи клетки.
Читайте также: