Анатомия: Базальные ядра полушарий. Полосатое тело, coprus striatum. Хвостатое ядро, nucleus caudatus.
Добавил пользователь Morpheus Обновлено: 28.01.2026
К ним относятся хвостатое ядро, чечевицеобразное, ограда и миндалевидное ядро. Между ядрами расположены капсулы белого вещества (рис. 32, рис. 33). Первые три из перечисленных ядер относятся к полосатому телу (corpus striatum). Они получают топографически упорядоченные проекции от всех полей коры и через таламус оказывают влияние на обширные фронтальные области. Таким образом, полосатое тело обеспечивает подготовку движений, а моторная кора - их точность и экономичность.
ХВОСТАТОЕ ЯДРО (nuklei caudatus) лежит вперед головкой, которая образует наружную стенку переднего рога бокового желудочка. Суживаясь кзади, головка переходит в тело, а затем в хвост, который достигает миндалевидное ядро, расположенное в височном полюсе (рис. 32, рис. 34).
ЧЕЧЕВИЦЕОБРАЗНОЕ ЯДРО (nukleus lentiformis) по форме сходно с чечевичным зерном. Оно отделено от таламуса внутренней капсулой, а спереди связано с хвостатым ядром. Небольшие прослойки белого вещества делят его на три ядра: скорлупу, медиальный и латералный бледный шар (рис. 34). Головка хвостатого ядра и скорлупа являются филогенетически более новыми образованиями, относятся к neostriatum. В их структуре различают многочисленные пятна - "стриосомы", которые функционально связаны с лимбической системой. Между "стриосоами" находится так называемый "матрикс", состоящий преимущественно из приходящих волокон и связан с экстрапирамидной моторной системой.
БЛЕДНЫЙ ШАР (globus pallidus) является филогенетически более старым образованием (paleostriatum). Своим углом оно обращено к колену внутренней капсулы (рис. 33), имеет более светлую окраску, чем скорлупа. Его дорсальная часть вовлечена в "экстрапирамидный моторный цикл" управления позой и инициации движений. хикъ
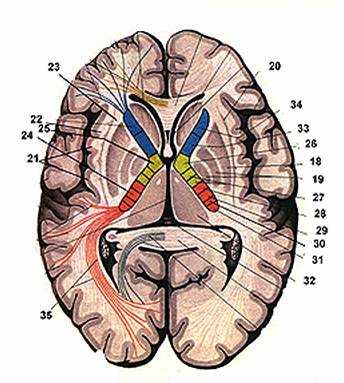
Рис. 32. Базальные ядра
1. таламус
2. концевая полоска
3. III желудочек
4. лобный рог I желудочка
5. височный рог Iж.
6. затылочный рог Iж.
7. сосудистое сплетение
8. гиппокамп
9. бахромка
10. зубчатая извилина
11. головка хвостатого ядра
12. хвост
13. тело
14. столбы свода
15. передняя спайка
16. прозрачные перегородки
17. полость прозрачной перегородки
Рис. 33. Базальные ядра и капсулы полушария (горизонтальный срез)
18. скорлупа
19. бледные шары
20. ограда
21. кора островка
22. самая наружная капсула
23. наружная капсула
24. внутренняя капсула:
25. колено
26. корково-ядерный путь
27. корково-спинномозговой
28. корково-красноядерный
29. височно-теменно-затылочный
30. слуховой
31. зрительный
32. таламус
33. лобно-мостовой
34. передняя таламическая лучистость
35. затылочная лучистость
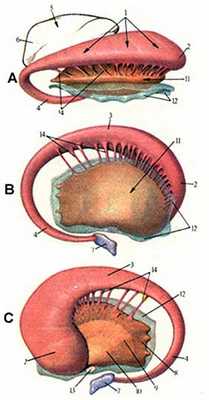
Рис. 34. Базальные ядра конечного мозга (полусхематично)
А -- вид сверху
B -- вид изнутри
C -- вид снаружи
1. хвостатое ядро
2. головка
3. тело
4. хвост
5. таламус
6. подушка таламуса
7. миндалевидное ядро
8. скорлупа
9. наружный бледный шар
10. внутренний бледный шар
11. чечевицеобразное ядро
12. ограда
13. передняя спайка мозга
14. перемычки
ОГРАДА (claustrum) - тонкая пластинка серого вещества, расположена латеральнее от скорлупы и отделена от нее наружной капсулой. По своему происхождению является как бы частью коры. В эту структуру входят волокна из амигдалоидного комплекса концевой полоски, поясной извилины, передней спайки. Свои волокна ограда направляет в ядра переднего продырявленного вещества, дорсомедиального таламуса и латеральную часть миндалевидного тела (рис. 33, рис. 34).
МИНДАЛЕВИДНОЕ ТЕЛО (corpus amigdoloideum), располагается в толще височного полюса. Различают базально-латеральную часть- это большая группа ядер, имеющих отношение к формированию памяти, интеграции вегетативных реакций при стрессе и др.
Корково-медиальная (обонятельная часть), расположена в верхнемедиальнойобласти миндалевидного тела, получает волокна от обонятельного тракта и принимает участие в формировании концевой полоски, связана с сексуальными запахами и половым поведением. Переднее миндалевидное поле расположено вблизи переднего продырявленного вещества, здесь заканчивается латеральный обонятельный тракт и начинается диагональная полоска Брока (рис. 30) активирует реакции защиты, страха и агрессии. Таким образом, миндалевидное тело оказывает влияние на некоторые вегетативные функции и эмоциональное поведение человека.
Норма базальные ганглии
База́льные я́дра (также база́льные га́нглии, лат. nuclei basales) — несколько скоплений серого вещества, расположенных в белом веществе латеральнее таламуса на уровне основания полушарий конечного мозга. Базальные ядра входят в состав переднего мозга, расположенного на границе между лобными долями и над стволом мозга. Традиционно в состав базальных ядер включались полосатое тело (лат. corpus striatum), в свою очередь состоящее из хвостатого ядра (лат. nucleus caudatus), скорлупы (лат. putamen) и бледного шара (лат. globus pallidus), а также ограда (лат. claustrum) и миндалевидное тело (лат. corpus amygdaloideum). Бледный шар и скорлупа вместе называются чечевицеобразным ядром (лат. nucleus lentiformis). Белое вещество между таламусом и чечевицеобразным ядром называется внутренней капсулой (лат. capsula interna), между чечевицеобразным ядром и оградой — наружной капсулой (лат. capsula externa) и между оградой и островком — самой наружной капсулой (лат. capsula extrema). Эта классификация основана на топографии анатомического среза мозга, однако в последнее время она всё чаще заменяется функциональной, где под термином «базальные ядра» понимают полосатое тело и несколько ядер промежуточного и среднего мозга (субталамическое ядро (лат. nucleus subtalamicus), чёрная субстанция (лат. substantia nigra) и ножкомостовое ядро покрышки (лат. nucleus tegmentalis peduncolopontinus)), которые совместно обеспечивают функциональную регуляцию движений и мотивационных аспектов поведения. Функции ограды остаются недостаточно изученными, а структуры миндалевидного тела относят к лимбической системе.
Все базальные ядра функционально объединены в две системы. Первая группа ядер представляют стриопаллидарную систему. К ним относятся хвостатое ядро, скорлупа и бледный шар. Скорлупа и хвостатое ядро имеют слоистую структуру и поэтому объединяются под названием «стриатум». Бледный шар светлее стриатума и не имеет слоистой структуры. Скорлупа и бледный шар объединены в чечевицеобразное ядро. Скорлупа образует наружный слой чечевицеобразного ядра, а бледный шар — внутренние его части. Бледный шар, в свою очередь, состоит из наружного и внутреннего чехликов. Ограда и миндалевидное тело входят в лимбическую систему мозга.
Базальные ганглии обычно изоденсивные или изотенсивные по отношению к коре головного мозга. Бледный шар содержит больше миелина по сравнению со скорлупой, поэтому бледное ядро визуализируется более гипоинтенсивным на T2WI, GRE и SWI изображениях. При старении в бледном шаре откладывается кальций, что приводит к снижению сигнала, а при кальцификации более, чем на 40% сигнал теряется на всех последовательностях. При старении также в скорлупе откладывается железо, что приводит к повышению сигнала на Т2. Данное явление часто выявляется у пациентов в возрасте от 70 до 80 лет.
БАЗАЛЬНЫЕ ЯДРА БОЛЬШИХ ПОЛУШАРИЙ, СТРУКТУРА И РАСПОЛОЖЕНИЕ (ХВОСТАТОЕ ЯДРО, ОГРАДА, МИНДАЛИНА)
I пара — обонятельные нервы (nn. olfactorii). Они начинаются от слизистой оболочки обонятельной области полости носа, проходят через решетчатую пластинку в полость черепа и подходят к обонятельной луковице, где оканчивается 1-й нейрон обонятельного пути и берет начало центральный обонятельный путь. Функция - отвечает за обонятельную чувствительность.
II пара — зрительный нерв (n. opticus). Через зрительный канал нерв проникает в полость черепа. Кпереди от турецкого седла оба нерва образуют зрительный перекрест (chiasma opticum), где волокна из медиальных (назальных) половин сетчаток переходят на противоположную сторону. После перекреста образуется зрительный тракт (tractus opticus), который огибает ножку мозга и отдает свои волокна подкорковым зрительным центрам. Функция - воспринятые чувствительными клетками сетчатки, передаются в головной мозг.
III пара — глазодвигательный нерв (n. oculomotorius). Он берет начало от двигательных ядер, расположенных в покрышке среднего мозга на уровне верхних холмиков. Нерв выходит в межножковой ямке из медиальной поверхности ножки мозга, входит в боковую стенку пещеристого синуса и через верхнюю глазничную щель попадает в глазницу. Здесь он делится на верхнюю и нижнюю ветви. Верхняя ветвь входит в мышцу, поднимающую верхнее веко, и в верхнюю прямую мышцу глазного яблока, а нижняя ветвь иннервирует нижнюю и медиальную прямые и нижнюю косую мышцы. Функция - отвечающий за движение глазного яблока, поднятие века, реакцию зрачков на свет.
IV пара — блоковый нерв (n. trochlearis), начинается от ядра, лежащего в покрышке среднего мозга на уровне нижних холмиков, выходит на задней поверхности мозгового ствола, огибает ножку мозга, идет в стенке пещеристого синуса и через верхнюю глазничную щель проникает в глазницу, иннервирует верхнюю косую мышцу глазного яблока. Функция - иннервируя верхнюю косую мышцу (лат. m.obliquus superior), которая поворачивает глазное яблоко кнаружи и вниз.
VI пара — отводящий нерв (n. abducens), в верхнем отделе ромбовидной ямки, выходит из мозга между краем моста и пирамидой продолговатого мозга, проходит через пещеристый синус к верхней глазничной щели, иннервирует наружную прямую мышцу глаза. Функция - иннервируя латеральную прямую мышцу (лат. m. rectus lateralis) отвечает за отведение глазного яблока
XI, XII ПАРЫ ЧЕРЕПНЫХ НЕРВОВ, ИХ ЯДРА, ТОПОГРАФИЯ И ОБЛАСТИ ИННЕРВАЦИИ
XI пара — добавочный нерв (n. accessorius), который начинается от двигательного ядра, расположенного в нижней части продолговатого мозга и первом-четвертом шейных сегментах спинного мозга. Соответственно он имеет черепные и спинномозговые корешки, которые объединяются в ствол нерва. Последний проходит через яремное отверстие и делится на внутреннюю и наружную ветви. Внутренняя ветвь присоединяется к блуждающему нерву, она содержит волокна, участвующие в двигательной иннервации глотки и гортани. Наружная ветвь снабжает грудино-ключично-сосцевидную и трапециевидную мышцы; она часто соединяется с шейным сплетением.
Иннервация - содержит двигательные нервные волокна, иннервирующие мышцы, ответственные за повороты головы, приподнимание плеча и приведение лопатки к позвоночнику.
XII пара — подъязычный нерв (n. hypoglossus), являющийся двигательным нервом языка. Его ядро лежит в нижнемедиальном участке ромбовидной ямки. Корешки подъязычного нерва выходят из продолговатого мозга между пирамидой и оливой. Из полости черепа нерв проходит через подъязычный канал затылочной кости, располагается на шее позади заднего брюшка двубрюшной и шилоподъязычной мышц, пересекает снаружи наружную сонную артерию и входит в мускулатуру языка, где разделяется на свои конечные ветви. Подъязычный нерв отдает соединительную ветвь к шейному сплетению, которая принимает участие в формировании шейной петли (ansa cervicalis).
Область иннервации - Отвечает за движение языка.
АППАРАТЫ СПИННОГО МОЗГА
Сегментарным аппаратом спинного мозга является серое его вещество с соответствующими передними и задними корешками. Сегмент спинного мозга входит в состав метамера тела, к которому относится также определенный участок кожи (дерматом), мышцы (миотом), кости (склеротом) и внутренностей (спланхнотом), иннервируемый этим сегментом.
Спинной мозг имеет 31—32 сегмента и делится на 5 отделов:
шейный (pars cervicalis) — C1—C8, грудной (pars thoracica) — Th1—Th5, поясничный (pars lumbalis) — L4—L8 и крестцовый (pars sacralis) — S1—S5. В самом низу имеются один-два рудиментарных копчиковых сегмента (Co1—Со2).
В обеих половинах каждого сегмента серого вещества имеются передний и задний рог. На наружной стороне передних рогов нижнешейного, грудного и верхнепоясничного отделов выделяется выпячивание, именуемое боковым рогом (cornu laterale)
Серое вещество состоит из нервных клеток, глиальных элементов и нервных волокон
В передних канатиках имеются следующие пути:
1. передний корково-спинномозговой (пирамидный) путь—от коры головного мозга к двигательным клеткам переднего рога, образован меньшей, неперекрещенной частью пучка пирамидного;
2. преддверно-спинномозговой путь (tr. vestibulospinalis) — от вестибулярных ядер моста к двигательным клеткам передних рогов;
3. покрышечно-спинномозговой путь (tr. tectospinalis) — от покрышки моста к клеткам передних рогов;
4. передний ретикулярно-спинномозговой путь — от ретикулярной формации мозгового ствола к клеткам передних рогов,
5. задний продольный пучок — от клеток различных ядер мозгового ствола к двигательным клеткам передних рогов;
6. передний спинно-таламический путь (tr. spinothalamicus ventralis anterior) — от клеток заднего рога к таламусу;
7. передние собственные пучки (fasc. proprii ventrales anteriores), относящиеся к собственному аппарату спинного мозга
БАЗАЛЬНЫЕ ЯДРА БОЛЬШИХ ПОЛУШАРИЙ, СТРУКТУРА И РАСПОЛОЖЕНИЕ (ХВОСТАТОЕ ЯДРО, ОГРАДА, МИНДАЛИНА)
БАЗАЛЬНЫЕ ЯДРА (nuclei basalis), подкорковые ядра, базальные ганглии, скопление серого вещества больших полушарий головного мозга позвоночных, участвующие в координации двигательной активности и формирования эмоциональных реакций. Базальные ядра вместе с корой мозга составляют клеточное вещество конечного мозга. Состоят из хвостатого ядра, скорлупы (объединяются в полосатое тело), бледного шара (объединяются со скорлупой в чечевицеобразное ядро), ограды, миндалевидного тела.
Хвостатое ядро (лат. nucleus caudatus) — парная структура головного мозга, относящаяся к стриатуму. Расположена спереди от таламуса, от которого (на горизонтальном срезе) его отделяет белая полоска вещества — внутренняя капсула. Передний отдел хвостатого ядра утолщён и образует головку, caput nuclei caudati, которая составляет латеральную стенку переднего рога бокового желудочка. Головка хвостатого ядра примыкает внизу к переднему продырявленному веществу, в этом месте головка соединяется с чечевицеобразным ядром. Суживаясь сзади, головка переходит в тело, corpus nuclei caudati, которое лежит в области дна центральной части бокового желудочка и отделяется от таламуса терминальной полоской белого вещества. Сзади продолжается в хвост — cauda.
Ограда (claustrum) - тонкая вертикальная прослойка серого вещества, залегающая в белом веществе каждого полушария большого мозга, расположенная между корой островковой доли полушария большого мозга и скорлупой чечевицеобразного ядра.
Миндалевидное тело, миндалина (лат. corpus amygdaloideum) — характерная область мозга, имеющая форму миндалины, расположенной внутри височной доли (Lobus temporalis) головного мозга. В мозге две миндалины — по одной в каждом полушарии. Миндалина играет ключевую роль в формировании эмоций, является частью лимбической системы.
Тема 7. Базальные ядра. Обонятельный мозг. Лимбическая система. Белое вещество полушарий.
Цель занятия: Дать общую характеристику базальным ядрам и белому веществу полушарий мозга.
Основные теоретические вопросы:
1. Строение компонентов стрио-паллидарной системы.
2. Функции базальных ядер.
3. Строение экстрапирамидной системы. Ее функциональное назначение.
4. Центральные и периферические отделы обонятельного мозга. Их функции.
5. Общая характеристика лимбической системы.
6. Строение и функции ассоциативных, комиссуральных и проекционыых волокон, их проводниковый состав.
ИНФОРМАЦИОННЫЙ БЛОК.
Базальные ядра полушарий.
К базальным ядрам (рис. 17.) относятся скопления серого вещества в глубине белого вещества полушарий: хвостатое ядро (nucleus caudatus) (1), чечевицеообразное ядро (nucleus lentiformis), ограда (claustrum) (9) и миндалевидное тело (corpus amygdoloideum). Чечевицеобразное ядро подразделяется на 3 части: 2 медиальные - бледный шар (globus pallidus) (5) и латеральную - скорлупу (putamen) (4).
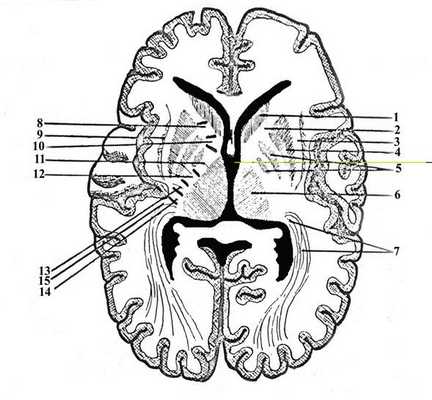
Рис. 17. Строение базальных ядер и внутренней капсулы.
1. Caput nuclei caudate (головка хвостатого ядра)
2. Capsula interna (внутренняя капсула)
3. Capsula externa (наружная капсула)
4. Putamen (скарлупа)
5. Globus pallidus (бледный шар)
6. Thalamus (таламус)
7. Radiatio optica (зрительная лучистость)
8. Tractus frontopontinus (лобно-мостовой тракт)
9. Claustrum (ограда)
10. Tractus corticonuclearis (корково-ядерный тракт)
11. Tractus corticospinalis (корково-спиномозговой тракт)
12. Tractus thalamocorticalis (таламо-кортикальный тракт)
13. Tractus occipitotemporopontinus (затылочно-височно-мостовой тракт)
14. Tractus cochlearis centralis (центральный слуховой тракт)
15. Tractus opticus centralis (центральный зрительный тракт)
Хвостатое ядро и скорлупа чечевицеобразного ядра объединяются под названием полосатого тела (corpus striatum), являющегося высшим центром экстрапирамидной системы, которому подчинена паллидарная система, включающая бледный шар, красное ядро, черная субстанция и подталамическое (Льюисово) или миндалевидное тело. Считается, что стрио-паллидарная система функционирует взаимоуровновешивая друг друга. При этом паллидарная система оказывает возбуждающее действие, а стриатум - тормозящее.
При чрезмерном тормозном влиянии стриарной системы возникает гипокинезия - бедность движений и мимики (гипомимия), гипофункция стриарной системы ведет к появлению избыточных непроизвольных движений - гиперкинезов, так как отсутствует тормозное влияние на паллидарную систему.
Основными путями, по которым проводятся импульсы к стрио-паллидарным структурам, являются проводники от зрительного бугра. По ним устанавливаются не только связи экстрапирамидной системы со зрительным бугром, но и с корой головного мозга.
Этим путем происходит включение экстрапирамидных аппаратов в систему произвольных, корковых движений.
Полосатое тело тесно связано с бледным шаром. Нисходящие пути направляются к черной субстанции, ретикулярной формации, красному ядру, ядру Даркшевича, четверохолмию, оливам. От них импульсы следуют к передним рогам спинного мозга, а затем к рабочему органу.
Подкорковые, базальные ядра полушарий являются элементами экстрапирамидной системы, они участвуют в координации двигательных актов. Считается, что информация от двигательных ядер коры конечного мозга, мест, где зарождается замысел движения, одновременно поступает к мозжечку и базальным ядрам; от них через зрительный бугор к двигательной коре, а оттуда, уже уточненная - через пирамидные и экстрапирамидные пути в спинной мозг.
В самом общем виде экстрапирамидная система обеспечивает моторно-тоническую функцию.
К экстрапирамидной системе относятся: хвостатое и чечевицеобразное ядро, ограда, зрительный бугор, миндалевидное (Льюисово) тело, черная субстанция, красное ядро, ретикулярная формация, ядра нижней оливы, мозжечок и волокна белого вещества, осуществляющие связь между всеми этими структурами.
Экстрапирамидная система осуществляет высшие безусловные рефлекторные акты, без участия сознания регулирует работу мускулатуры, обеспечивая непроизвольные автоматические движения, поддерживает тонус мускулатуры и перераспределяет его при движениях.
Участок белого вещества, расположенный между зрительным бугром и головкой хвостатого ядра с медиальной стороны и чечевицеобразным ядром с латеральной, называется внутренней капсулой (capsula interna). В ней различают переднее и заднее бедро, соединенное коленом.
В переднем бедре внутренней капсулы проходят следующие проводящие пути:
1. корково-стриарный путь (tractus corticostriatus), связывающий кору головного мозга со структурами стрио-паллидарной системы;
2. лобно-мостовой путь (tractus frontopontinus) (8) идущий из коры лобной доли к собственным ядрам моста своей стороны.
В колене внутренней капсулы расположен корково-ядерный путь (tractus corticonuclearis) (10), который связывает кору передней центральной извилины конечного мозга с двигательными ядрами черепно-мозговых нервов.
В заднем бедре определяются следующие проводящие пути:
1. корково-спиномозговой путь (tractus corticospinalis) (11), связывающий кору головного мозга и передние рога спинного мозга;
2. таламо-корковый путь (tractus thalamocorticalis) (12), связывающий таламус и кору головного мозга;
3. затылочно-височно-теменно-мостовой путь (tractus occipito-temporo-parieto-pontinus) (13), связывающий одноименные доли коры полушарий;
4. слуховая лучистость (radiatio acustica) (14) и зрительная лучистость (radiatio optica) (15),связывающие подкорковые и корковые центры слуха и зрения.
Базальные ядра: функции, норма и патология

Для того, чтобы организм человека ежедневно выполнял сложнейшие двигательные программы, существуют специализированные структуры головного мозга, такие как мозжечок и базальные ядра, которые находятся в тесной связи с областями коры больших полушарий.
В то время, как мозжечок обеспечивает синхронизацию движений и их одномоментное соответствие требованиям, базальные ядра позволяют организму планировать сложные двигательные программы, а самое главное — их осуществлять. Помимо регуляции движений, подкорковые ядра участвуют в познавательной деятельности мозга, а значит и в формировании эмоций. Именно о них и пойдёт речь в данной статье.
Чтобы лучше понимать механизмы нормальных и патологических процессов, для начала стоит рассмотреть строение подкорковых ядер: их расположение и образование многочисленных связей с таламусом и корковыми областями. С анатомических позиций базальные ядра включают хвостатое ядро, скорлупу, бледный шар и ограду. Указанные четыре ядра составляют полосатое тело. Однако часто используемое понятие «стриатум» включает только два образования — скорлупу и хвостатое ядро. Данные образования располагаются в основном латеральнее таламуса и занимают большую часть внутренних регионов полушарий мозга.
Вся информация, которая поступает в базальные ядра в виде сигналов, распределяется независимо друг от друга по особым параллельным путям обработки информации. Эти пути образуют функциональные круги, которые также являются независимыми и включают в себя различные регионы коры. Поэтому специфическая роль ядер определяется именно той областью коры, которая находится в одном функциональном круге с ядром.
Двигательные возбуждающие сигналы комплекс базальных ганглиев принимает от премоторных зон коры, перерабатывает их и вновь возвращает в кору, но уже в первичную моторную область. «Замысел» движения возникает в премоторной коре, оттуда стриатум получает двигательные команды. Благодаря такой связи базальные ганглии способны «включать» двигательные поведенческие программы еще до их совершения. При этом сами программы в ганглиях уже заложены, и им предстоит только принять решение — к какому действию прибегнуть и прибегать ли вовсе. Выполнение сложных двигательных актов, запускаемых корой, обеспечивается с помощью прямого нервного пути. Его функция заключается в следующем: базальные ядра облегчают действия, задаваемые корой, и подавляют лишние сопутствующие.
Вся информация, которая поступает к базальным ядрам, собирается из нескольких регионов коры. Чувствительные волокна образуют возбуждающие глутаматергические синапсы с нейронами стриатума, которые объединяются в функциональные модули, перерабатывающие информацию однотипными механизмами. Поскольку моторные и сенсорные волокна различаются по характеру информации, передающейся ими с помощью импульса, то и модули, к которым направляются волокна, будут различны. По этой причине пучки волокон, направляющихся к разным модулям, образуют отдельные полоски, из-за которых стриатум получил своё название (лат. corpus striatum — полосатое тело).
Нейроны стриатума в свою очередь образуют ГАМК-ергические синапсы с клетками внутреннего сегмента бледного шара и части черной субстанции (первое последовательное тормозное переключение). Нейроны данных структур образуют тормозные ГАМК-ергические синапсы на переднем и вентральном ядрах таламуса (второе последовательное тормозное переключение), что приводит к постоянному торможению прохождения возбуждения от таламуса к коре. Отростки нейронов ядер таламуса, которые направляются к коре, образуют главные эфферентные глутаматергические пути. Поэтому при угнетении внутреннего сегмента бледного шара и части черной субстанции возбуждающее действие таламуса на кору усиливается, что обеспечивает облегчение выполнения движения.
Поскольку передача информации между структурами головного мозга обеспечивается химическими рецепторами, вопрос о постоянном синтезе медиатора является особенно важным. Роль поставщика биологически активных веществ в данном случае играет черная субстанция, получившая свое название из-за пигмента — нейромеланина, который придает ей соответствующую окраску. Черная субстанция вырабатывает дофамин, который работает как возбуждающий нейромедиатор и также служит важной частью «системы поощрения» мозга.
На нейронах прямого пути локализованы возбуждающие дофаминовые рецепторы типа Д1, благодаря чему химические сигналы от черной ситуации возбуждают этот путь. От того, в каком соотношении будут находиться концентрации дофамина и глутамата, зависят фазический и тонический компоненты движений человека. При преобладании дофамина происходит усиление фазического, быстрого компонента движения, при преобладании глутамата, ацетилхолина и ГАМК — тонического , медленного компонента. Вместе с усилением фазического компонента возрастает и скорость движений, однако происходит снижение тонуса.
Для поддержания нормального функционирования двигательных систем необходимо соблюдение как анатомической целостности путей, так и поддержание определенного уровня медиатора. Соответственно, поражение структур или избыток/недостаток нейромедиатора влекут за собой серьезные последствия. При поражении базальных ядер возникают расстройства двигательной активности — дискинезии (гипокинезы или гиперкинезы) и изменения мышечного тонуса (гипотония или ригидность мышц). При функциональных нарушениях бледного шара наблюдаются спонтанные и, часто, постоянные волнообразные движения кисти, руки, шеи или лица. Такие движения называют атетозом.
Поражение субталамического ядра (также относится к базальным ганглиям) ведет к возникновению размашистых движений всей конечности. Такое состояние называют гемибаллизмом. Множественные мелкие поражения в скорлупе ведут к появлению быстрых подергиваний в кистях, лице и других частях тела, что называется хореей. Поражения черной субстанции ведут к распространенному и чрезвычайно тяжелому заболеванию, связанному с акинезией и тремором. Это заболевание известно как болезнь Паркинсона.
Основными клиническими проявлениями болезни Паркинсона являются гипокинезия и мышечная ригидность. Гипокинезия проявляется в очень медленном совершении активных двигательных действий: начало двигательного акта затруднено, отсутствует содружественное движение верхних конечностей — синкинезия, при ходьбе они неподвижны (ахейрокинез). Мышечная ригидность является своеобразным сопротивлением пассивным движениям, появляется не только в начальной фазе движения, но и во всех последующих фазах растяжения мышц. Конечность как бы застывает в той позе, которую ей придают.
Помимо вышеописанных проявлений, также наблюдается маскообразное лицо — амимичное, с неподвижным взглядом, редким миганием, иногда отсутствующим в течение нескольких минут, бледной жестикуляцией. Гипокинезия и ригидность могут наблюдаться изолированно, но к ним нередко присоединяется гиперкинез в виде тремора пальцев кисти (по типу счета монет), подбородочной области и нижних конечностей. Несмотря на то, что тремор является одним из клинических признаков паркинсонизма, его патогенез так и остается неясным. Хотя с точки зрения неврологии, в отличие от других двигательных признаков, тремор имеет точные электрофизиологические характеристики (частота, фаза и мощность).
Целью различных клинических испытаний является нахождение взаимосвязи между тремором и дофамином. Однако было выяснено, что изменения характеристик тремора в ответ на действие дофаминергических препаратов достаточно вариабельны. Эти уникальные особенности тремора и новые методы нейровизуализации способствуют возникновению новых исследований в области изучения этой патологии.
В статье Helmich R.C., посвященной церебральным основам возникновения паркинсонического тремора, подробно рассматриваются гипотезы его возникновения вследствие усиления взаимодействия между подкорковыми ядрами и мозжечково-таламо-кортикальной цепью. Обычно усиление этих связей обуславливается увеличением количества дофаминергических рецепторов в ядрах из-за воздействия различных факторов (например, психологический стресс).
Также авторами статьи подробно рассматриваются модели, которые помогают понять патогенез тремора. Одним из примеров может служить «dimmer-switch» модель, согласно которой церебральная активность, связанная с тремором при паркинсонизме, сначала возникает в базальных ядрах, а затем усиливается и распространяется на мозжечково-таламо-кортикальный контур (распространение обеспечивается активацией возбуждающих синапсов).
Помимо всем известной функции базальных ядер, заключающейся в контроле двигательной активности, существуют также менее изученные, но не менее интересные «обязанности» у данных подкорковых структур.
В своей статье Lukas Maurer с коллегами предложил концепцию, описывающую нарушения «цепей мозга» для объяснения множества психоневрологических заболеваний. Заболевания характеризуются патологическими изменениями в структуре нейронных сетей, включая изменения в осцилляторной сигнализации корково-подкорковых цепей в системе базальных ядер. Часть этих цепей играет значительную роль в поддержании энергетического баланса организма. Поэтому статья посвящена взаимосвязи между ожирением и изменениями в осцилляторной сигнализации лимбических кортико-базальных цепей.
Таким образом, можно сделать вывод, что спонтанная бета-активность строго коррелирует с эндокринными показателями ожирения. Однако проба с глюкозой увеличивала бета-активность у тех животных, которые не соблюдали HFD. Причём интравентрикулярное введение инсулина также увеличивало активность NAC. Благодаря исследованию удалось подтвердить гипотезы о наличии корреляции между лимбической кортико-базальной петлей, ожирением и уровнем сывороточного инсулина. Поэтому можно рассматривать резистентность к инсулину и ожирение как следствия осцилляторных нарушений в клетках лимбической кортико-базальной цепи.
Эта цепь играет центральную роль в «системе вознаграждения» мозга, которая отвечает за обработку информации, связанной с потреблением пищи и наградой. Кроме того, она неразрывно связана с гипоталамическими областями среднего мозга, которые регулируют гомеостатические функции организма. При сравнении двух пациентов, у индивидуума с ожирением наблюдалась повышенная активность кортико-базальных структур во время ожидания пищи и снижение этой активности при получении «награды». В соответствии с этим ожирение можно рассматривать как нервно-психическое расстройство, сходное с наркоманией, поскольку оно также основано на зависимости, только в данном случае — пищевой.
Рассмотрев две совершенно разные функции базальных ядер, можно сделать вывод о том, насколько сложна и запутанна природа любой структуры головного мозга. Но именно такая сложность будет и далее привлекать человека, стремящегося познать новое.
Читайте также:
- Преждевременное половое созревание: причины, симптомы и лечение
- Этапы и техника удаления подмышечных лимфоузлов (лимфаденэктомии)
- Расчет фракции фильтрации почек. Вычисление реабсорбции или секреции в канальцах
- Проявления заболеваний ЦНС. Мозговая опухоль
- Результаты хирургического лечения набухающей катаракты. Особенности