Экстрапирамидная двигательная система. Нейроны двигательной системы
Добавил пользователь Евгений Кузнецов Обновлено: 01.02.2026
Экстрапирамидная система (лат.: extra — вне, снаружи, в стороне + pyramis, греч.: πϋραμίς — пирамида) - совокупность подкорковых и стволовых образований, моторных путей, которые не проходят через пирамиды продолговатого мозга. Данная система, наряду с кортикальной, участвует в управлении движениями, причем, являясь наиболее филогенетически древней, играет значительную роль в построении и контроле движений, не требующих активации внимания.
Впервые мысль о том, что на состояние двигательных функций оказывает влияние не только образования, входящие в состав пирамидной системы, высказал английский невролог С. Вильсон в 1908г. в процессе изучения заболевания, известного теперь как болезнь Вильсона-Коновалова. С тех пор все структуры головного мозга, которые влияют на состояние поперечнополосатых мышц и участвующие в обеспечении движений стали именовать экстрапирамидными.
По предложению Р. Гранита (Granit R., 1973) структуры пирамидных путей, от которых зависят активные движения тела и его частей, были названы фазическими. Экстрапирамидные структуры, влияющие на двигательные акты, положение, поддержание равновесия тела и его позу - тоническими.
Н.К. Боголепов отмечал, что экстрапирамидная система участвует также в выполнении выразительных, мимических и эмоциональных реакций.
В 1973 г. ведущий американский физиолог П. Милнер (Milner P.) высказал сомнение относительно существования отдельных экстрапирамидной и пирамидной систем. Однако на данный момент, после тщательного изучения функций той и другой системы, их обособленность неоспорима.
Э. система по сравнении с пирамидной является филогенетически более древней (особенно ее паллидарная часть). С развитием пирамидной системы экстрапирамидная система переходит в соподчиненное положение.
Экстрапирамидная система состоит из следующих структур головного мозга:
базальные ганглии (включая полосатое тело), красное ядро, интерстициальное ядро, тектум, чёрная субстанция, ретикулярная формация ствола мозга, ядра вестибулярного комплекса, мозжечок, нижняя олива продолговатого мозга.
Nucleus basales - скопления серого вещества в толще полушарий большого мозга.
Corpus striatum ( полосатое тело ) состоит из n.caudatus и n.lentiformis.
Хвостатое ядро-имеет форму запятой, расположенной в сагиттальной плоскости. Состоит из трех частей: caput, corpus и cauda. Передняя часть головки сращена с белым веществом лобной доли, своими свободными поверхностями (верхней и медиальной) тело хвостатого ядра в височной доле образует дно центральной части бокового желудочка. Хвост направлен в височную долю полушария, где достигает миндалевидного тела. Участвует в организации двигательной активности, формировании условных рефлексов и механизмов памяти.
Чечевицеобразное ядро-залегает латерально от хвостатого ядра и зрительного бугра и состоит из двух частей - латеральной, большей по размеру и медиальной, скорлупы (putamen) и бледного шара (globus pallidus). Функцией чечевицеобразного ядра являются регуляция мышечного тонуса.
Подталамическое ядро (Льюиса) - скопление серого вещества с пронизывающими его волокнами. Прилегает к поверхности внутренней капсулы, отделяющей его от бледного шара, с которым ядро связано большим числом волокон, проходящих в составе субталамического пучка.
Черное вещество- ядро э.с., лежащее в основании ножек мозга, участвует в распределении мышечного тонуса, необходимого для установки тела в определенном положении.
Красное ядро - серое вещество покрышки среднего мозга, является первым интегративным центром управления конечностями.
Промежуточное ядро ( Кахаля ) - содержится в ретикулярной формации среднего мозга, дает начало медиальному продольному пучку, который является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Пластинка четверохолмия - часть среднего мозга, являющаяся рефлекторным центром различного рода движений, возникающих, г.о. под влиянием зрительных и слуховых раздражений.
Нижняя олива- овальное возвышение, находящееся по бокам от каждой пирамиды продолговатого мозга. Отделена от последней переднелатеральной бороздой. Играет важную роль в желудочно-ободочном рефлексе, проводит волокна из медиального спинно-мозжечкового тракта
Cerebellum - часть заднего мозга, отвечающий за регуляцию позы и мышечного тонуса, сенсомоторную координацию позных и целенаправленных движений, координацию быстрых целенаправленных движений.
Часть структур э.с. участвует в формировании ее высшего отдела, так называемой стриопаллидарной системы, состоящей из pallidum (более филогенетически древнего образования, состоящего из бледного шара, ядра Льюиса, красного ядра, черного вещества) и striatum (более молодой части, состоящей из скорлупы и ограды).
Несмотря на объединение стриатума и паллидума в единую систему, в функциональном плане эти структуры отличаются друг от друга. Паллидум представляет собой двигательное ядро, которое оказывает активизирующее воздействие на подкорковые образования. Бледный шар получает афферентные импульсы по волокнам, идущим от стриатума и субталамического ядра. Часть волокон следуют к таламическим ядрам, нейронам красного и покрышечных ядер. Бледный шар, будучи связан эффекторно с центрами среднего и заднего мозга, регулирует и координирует их работу. Одной из функций паллидума считают торможение нижележащих ядер, главным образом красного ядра среднего мозга, поэтому при повреждении бледного шара наблюдается сильное увеличение тонуса скелетной мускулатуры - гипертонус вследствие освобождения красного ядра от тормозящего влияния бледного шара.
Стриатум же оказывает тормозящее действие на подкорковые структуры. Стриатум получает афферентные импульсы главным образом от больших полушарий и таламуса и посылает эфферентные импульсы главным образом к бледному шару. Часть эфферентов не переключаясь идет к черной субстанции. Незначительное число волокон направляется к субталамическому и красному ядру, нижней оливе, голубому пятну, ядрам шва. Полосатое тело рассматривают как эффекторное ядро, не имеющее самостоятельных двигательных функций, но контролирующее функции паллидума. Полосатое тело тормозит деятельность бледного шара, то есть действует на него таким же образом, как он сам действует на красное ядро.
-nucleus basales (corpus striatum, n,amygdaloideum)
Акинетико-ригидный синдром (гипомимия,
Баллизм и гемибаллизм
-substantia grisea, nucleus ruber, n.intersticieles,lamina tecti,
Гипотонически-гиперкинетический синдром (гиперкинезы, атетоз)
-oliva inferior, nucleus vestibularis
Formation reticularis trunci encephali
Функционирование э.с. осуществляется с помощью многочисленных специализированных проводящих путей:
Афферентные корковые пути берут свое начало от многочисленных отделов коры больших полушарий, особенно из двигательных зон лобной области (предцентральная извилина, парацентральная долька). Вероятнее всего данные волокна являются тормозными. Другая система афферентных волокон, которая, по-видимому является активирующей, достигает полосатого тела из таламического центромедианного ядра. Кроме этого выделяется система корковых волокон, обеспечивающих двусторонние связи между черной субстанцией и стриатумом. Дофаминэргические (афферентные нигростициальные) волокна - уменьшают тормозную функцию стриатума (аксонами нейронов этих путей выделяется около 80% дофамина, (нигростриарная система, аксонами нейронов которой выделяется около 80% дофамина- нейромедиатора в окончаниях некоторых аксонов периферических нервов и многих нейронов ЦНС. Дофамин является одним из химических факторов внутреннего подкрепления (ФВП) и служит важной частью «системы поощрения» мозга, поскольку вызывает чувство удовольствия (или удовлетворения), чем влияет на процессы мотивации и обучения ).С другой стороны, стрионигральный путь является ГАМК-эргическим и оказывает ингибирующее воздействие на дофаминэргические нейроны.
Эфферентные tractus rubrospinalis, tractus tectospinalis, tractus vestibulospinalis , tractus reticulospinalis , tractus olivispinalis.
Рубро-спинальный (путь Монакова) тракт начинается от красного ядра среднего мозга, после выхода этот путь вновь перекрещивается (перекрёст Фореля) расположен в боковых канатиках и оканчивается в V-VII пластинах спинного мозга (промежуточная зона). Прямых связей с мотонейронами нет. Участвует в организации движения всей мускулатуры туловища и конечностей (ходьба, бег), контролирует тонус сгибателей, делая эти движения пластичными, способствуя сохранению определенной позы на протяжении длительного времени.
Вестибуло-спинальные тракты (латеральный и вентральный) активируют моторные системы, связанные с равновесием тела и направлением движения. Они начинаются от крупных вестибулярных ядер продолговатого мозга и моста, оливы продолговатого мозга и оканчивается в VII - IX пластинах спинного мозга (область переднего рога). Латеральный тракт (проходящий в боковом канатике) заканчивается в области шейных отделов, а вентральный (проходящий в переднем канатике) в поясничных отделах. Контролируют и организуют перераспределение мышечного тонуса при любом изменении положения тела и головы в пространстве, контролируют тонус разгибателей.
Ретикуло-спинальные тракты представлены аксонами нейронов ретикулярной формации моста и продолговатого мозга. Они проходят в переднем канатике, образуя два пути: медиальный и латеральный, и заканчиваются в области моторных ядер IX пластины, а также в области VI - VIII пластин. Важное значение этого тракта в механизме поддержания и распределения мышечного тонуса и регуляции поз, участие в диффузном торможении всей мускулатуры необходимо в некоторых поведенческих актах, например, при перегревании организма. Этот путь опосредует активирующее влияние дыхательного и сосудистодвигательного центров.
Текто-спиннальный тракт берет начало в глубоких слоях бугров четверохолмия среднего мозга, проходит в составе вентральных канатиков и оканчивается на интернейронах VII — VIII пластин спинного мозга (область переднего рога). Функционально текто-спинальные пути связаны с координацией движений головы и туловища в ответ на зрительные и звуковые сигналы и обеспечивают осуществления сторожевого рефлекса у человека.
Оливо-спиннальный тракт берет начало от нижнего ядра оливы продолговатого мозга, оно имеет непосредственные связи с корой полушарий лобной доли(корково-оливный путь), с красным ядром(красноядерно-оливный путь) и корой полушарий мозжечка(оливо-мозжечковый путь). Аксоны клеток нижнего ядра собираются в пучок, который прослеживается в передне-медиальном отделе бокового канатика и заканчивается в двигательных ядрах передних рогов спинного мозга. Обеспечивает безусловно-рефлекторное поддержание тонуса мышц шеи, головы и двигательные акты, направленные на сохранение равновесия тела.
Импульсы из экстрапирамидной системы, равно как и из мозжечка и из пирамидной системы, притекают, следовательно, к клеткам переднего рога, где и заканчиваются все только что перечисленные проводники. Окончательный путь к мышце проходит через периферический двигательный нейрон.
Ассоциативные (связи нейронов экстрапирамидной системы между собой)
Экстрапирамидные расстройства можно разделить на два клинических синдрома: акинетико-ригидный и гиперкинетико-гипотонический
Вариантами снижения двигательной активности являются: акинезия - отсутствие движений, брадикинезия - замедленность движений, олигокинезия -бедность движений, гипокинезия - недостаточность двигательной активности.
Ригидность - постоянное пребывание мышц в состоянии тонического напряжения, которое свойственно как мышцамагонистам, так и мышцам-антагонистам, в связи с чем и проявляется пластический характер повышения мышечного тонуса.
При акинетико-ригидном синдроме в начальной стадии его развития мышечная ригидность при болезни Паркинсона обычно асимметрична, может проявляться в какой-либо одной части тела, однако в дальнейшем, по мере прогрессирования заболевания, она становится все более распространенной и со временем генерализованной.
Гипотонически-гиперкинетический синдром развивается при поражении хвостатого ядра и скорлупы (в этих образованиях отмечается избыток медиаторов возбуждения — дофамина и др.). Различают:
атетоз — медленные вычурные тонического характера движения, преимущественно в дистальных отделах конечностей, иногда и в мускулатуре лица (выпячивание губ и др)
хореический гиперкинез (наиболее распространенная форма) — быстрые неритмичные движения в дистальных и проксимальных отделах конечностей, лице, туловище.
Итак, функционально э.с. очень тесно связана с более филогенетически молодой, пирамидной. Но в тоже время ей присущ определенная роль, заключающаяся в 1. Регуляции мышечного тонуса в комплексе с другими структурами.
2. Регуляции темпа, ритма и пластики любого произвольного двигательного акта.
3. Обеспечении двигательного компонента в регуляции безусловных рефлексов (половой, оборонительный, старт-рефлекс и др.).
4. Обеспечении последовательности двигательного акта.
5. Обеспечении моторного компонента эмоциональной сферы.
6. Регуляции высокоспециализированных движений человека, которые достигли уровня автоматизмов
Бурд, Гусев, Коновалов - Неврология и нейрохирургия
Общая неврология А.С. Никифоров, Е.И. Гусев - 2007г. - 720 с
Пневмапсихасоматология человека, Трифонов Е.В. Русско-английская энциклопедия 2013
Пирамидная и экстрапирамидная системы регуляции движений
В отношении контроля движений часто выделяют две системы: пирамидную и экстрапирамидную. К пирамидной системе относятся двигательные области коры б.п. и начинающийся от них пирамидный (или кортико-спинальный) тракт спинного мозга. Эта система отвечает за контроль произвольных осознаваемых движений с использованием зрительных, тактильных и других сенсорных сигналов.
Любое произвольное движение формируется в три этапа.
- 1. Выбор программы (лобная ассоциативная кора).
- 2. Разбиение программы на отдельные движения (премоторная кора, поле 6 по Бродману).
- 3. Запуск сокращения конкретных мышц (двигательная кора, поле 4 по Бродману).
В моторной (двигательной) коре есть представительство всех мышц тела (см. гл. 15), и каждый нейрон управляет своей группой мышечных клеток при помощи сигналов, идущих по пирамидным трактам. Также в премоторной коре начинаются и волокна, идущие к структурам экстрапирамидной системы.
Экстрапирамидная система включает в себя премоторную область коры, стриопаллидариую систему, черную субстанцию, красное ядро, ретикулярную формацию ствола мозга, вестибулярные ядра, мозжечок и ряд других образований. С эволюционной точки зрения это более древняя система по сравнению с пирамидной. Ее структуры участвуют в обеспечении непроизвольных движений, не требующих контроля сознания и внимания, в поддержании мышечного тонуса и позы. У каждой структуры этой системы свои характерные функции. Так, если стриопаллидарная система отвечает преимущественно за запоминание программы движений в целом (последовательности движений), то мозжечок — за запоминание параметров конкретных, отдельных движений, реализующихся в общей программе.
Некоторые двигательные расстройства, возникающие в результате поражения структур экстрапирамидной системы (проявления паркинсонизма, гинеркинезы, атетозы, тремор и т.д.), так и называют — экстрапирамидные расстройства.
Экстрапирамидные пути
1. Красноядерно-спинномозговой путь. Красное ядро является основным двигательным координационным центром экстрапирамидной системы. Оно имеет многочисленные связи с корой полушарий большого мозга, со стриопаллидарной системой, с таламусом, с подталамической областью и с мозжечком. Нервные импульсы, поступающие к нейронам красного ядра от коры полушарий, ядер стриопаллидарной системы и ядер промежуточного мозга, после соответствующей обработки следуют по красноядерно-спинномозговому пути, обеспечивающему выполнение сложных привычных движений (ходьба, бег), делая эти движения пластичными, способствуя сохранению определенной позы на протяжении длительного времени, а также обусловливая поддержание тонуса скелетной мускулатуры.
От нейронов полушарий большого мозга, преимущественно из лобной доли, аксоны формируют корково-стриарный тракт, который проходит через переднюю ножку внутренней капсулы. Лишь небольшая часть волокон этого тракта заканчивается непосредственно на мелких мультиполярных клетках красного ядра среднего мозга. Бо́льшая часть волокон направляется к ядрам стриарной системы (базальным ядрам головного мозга), в частности к хвостатому ядру и скорлупе. От нейронов стриарной системы к красному ядру направляется стриарно-красноядерный путь.
Из структур промежуточного мозга с красным ядром связаны нейроны медиальных ядер таламуса (подкорковый чувствительный центр экстрапирамидной системы), нейроны бледного шара (паллидарная система) и нейроны задних ядер гипоталамуса. Аксоны клеток ядер промежуточного мозга собираются в таламо-красноядерный пучок, который заканчивается на клетках красного ядра и черного вещества. Нейроны черного вещества также имеют связи с красным ядром.
Нервные импульсы, поступающие к нейронам красного ядра из мозжечка, осуществляют так называемую поправочную деятельность. Они обеспечивают выполнение тонких целенаправленных движений и предотвращают инерционные проявления при движениях.
Мозжечок связан с красными ядрами посредством двухнейронного пути - мозжечково-красноядерного тракта. Первыми нейронами этого пути являются клетки коры полушарий мозжечка, аксоны которых заканчиваются в зубчатом ядре. Вторыми нейронами являются клетки зубчатого ядра, аксоны которых покидают мозжечок через верхние ножки. Мозжечково-красноядерный тракт входит в средний мозг, па уровне нижних холмиков перекрещивается с одноименным трактом противоположной стороны (перекрест Верне- кинга) и заканчивается на клетках красного ядра (рис. 4.10).
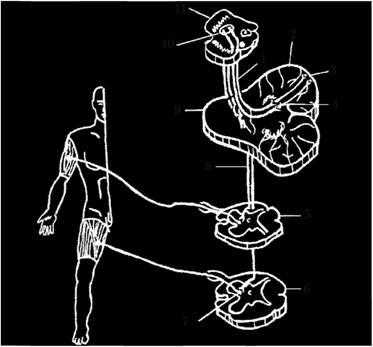
Рис. 4.10. Красноядерно-спинномозговой путь:
1 - зубчато-красноядерный путь; 2 - мозжечок; 3 - кора мозжечка; 4 - зубчатое ядро; 5 - шейный сегмент; 6 - поясничный сегмент; 7 - двигательные ядра передних рогов спинного мозга; 8 - красноядерно-спинномозговой путь; 9 - мост; 10 - красное ядро; 11 - средний мозг
От нейронов каждого красного ядра начинается нисходящий красноядерно-спинномозговой путь (пучок Монакова) и красноядерно-ядерный путь, которые сразу же в покрышке среднего мозга переходят на противоположную сторону и образуют передний перекрест покрышки (перекрест Фореля).
Красноядерно-ядерный путь проходит в покрышке ствола головного мозга и заканчивается на мотонейронах двигательных ядер черепных нервов. Аксоны мотонейронов ядер черепных нервов направляются к скелетным мышцам глазного яблока, головы, глотки, гортани и верхней части пищевода, обеспечивая их эфферентную иннервацию.
Красноядерно-спинномозговой путь проходит в боковом канатике спинного мозга. В последнем он располагается кпереди от латерального корково-спинномозгового пути. Постепенно пучок волокон истончается, так как аксоны посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга своей стороны. Аксоны мотонейронов покидают спинной мозг в составе передних корешков спинномозговых нервов, а затем в составе самих нервов и их ветвей направляются к скелетным мышцам.
2. Крыше-спинномозговой путь осуществляет безусловнорефлекторные двигательные реакции в ответ па внезапные сильные зрительные, слуховые, тактильные и обонятельные раздражения. Первые нейроны крыше-спинномозгового пути располагаются в верхних холмиках среднего мозга - подкорковом интеграционном центре среднего мозга (рис. 4.11). В данный интеграционный центр информация поступает из подкорковых центров зрения (ядро верхнего холмика), подкоркового центра слуха (ядро нижнего холмика), подкоркового центра обоняния (ядро сосцевидного тела) и коллатералей от проводящих путей общей чувствительности (спинномозговая, медиальная и тройничная петли).
Аксоны первых нейронов направляются вентрально и кверху, обходят центральное серое вещество среднего мозга и переходят на противоположную сторону, образуя задний перекрест покрышки (перекрест Мейнерта). Далее тракт проходит в дорсальной части моста рядом с медиальным продольным пучком. По ходу тракта в стволе головного мозга отходят волокна, которые заканчиваются на мотонейронах двигательных ядер черепных нервов. Эти волокна объединяются под названием крыше-ядерного пучка. Они обеспечивают защитные реакции с участием мышц головы и шеи.
В области продолговатого мозга крыше-спинномозговой путь приближается к дорсальной поверхности пирамид и направляется в передний канатик спинного мозга. В спинном мозге он занимает самую медиальную часть переднего канатика, ограничивая переднюю срединную щель.
Крыше-спинномозговой путь прослеживается на протяжении всего спинного мозга. Постепенно истончаясь, он посегментно отдает ответвления к мотонейронам двигательных ядер передних рогов спинного мозга своей стороны. Аксоны мотонейронов проводят нервные импульсы к мускулатуре туловища и конечностей.
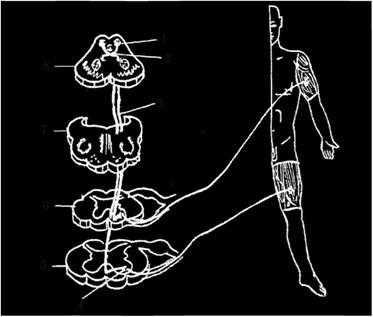
Рис. 4.11. Крыше-спинномозговой путь:
1 - верхний холмик среднего мозга; 2 - задний перекрест покрышки; 3 - крыше-спинномозговой путь; 4 - двигательные ядра передних рогов спинного мозга; 5 - поясничный сегмент; 6 - шейный сегмент; 7 - продолговатый мозг; 8 - средний мозг
При поражении крыше-спинномозгового тракта исчезают стартовые рефлексы, рефлексы на внезапные звуковые, слуховые, обонятельные и тактильные раздражения.
3. Ретикулярно-спинномозговой путь обеспечивает выполнение сложных рефлекторных актов (дыхательные, хватательные движения и т.д.), требующих одновременного участия многих групп скелетных мышц. Следовательно, он осуществляет координационную роль при этих движениях. Ретикулярно-спинномозговой путь проводит нервные импульсы, оказывающие активирующее или, наоборот, тормозное воздействие на мотонейроны двигательных ядер передних рогов спинного мозга. Кроме того, этот путь передает импульсы, обеспечивающие тонус скелетной мускулатуры.
Первые нейроны ретикулярно-спинномозгового пути располагаются в ретикулярной формации ствола головного мозга. Аксоны этих нейронов идут в нисходящем направлении. В спинном мозге они образуют пучок, который располагается в переднем канатике. Пучок хорошо выражен только в шейном и верхнегрудном отделах спинного мозга. Посегментно он истончается, отдавая волокна к гамма-мотонейронам двигательных ядер передних рогов спинного мозга. Аксоны этих нейронов направляются к скелетным мышцам.
- 4. Преддверно-спинномозговой путь обеспечивает безусловнорефлекторные двигательные акты при изменении положения тела в пространстве. Преддверно-спинномозговой путь образован аксонами клеток латерального и нижнего вестибулярных ядер (ядер Дейтерса и Роллера). В продолговатом мозге он располагается в дорсальном отделе. В спинном мозге проходит на границе бокового и переднего канатиков, поэтому пронизан горизонтально ориентированными волокнами передних корешков спинномозговых нервов. Волокна преддверно-спинномозгового пути посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга. Аксоны мотонейронов в составе корешков спинномозговых нервов покидают спинной мозг и направляются к скелетной мускулатуре, обеспечивая перераспределение тонуса мышц в ответ на изменение положения тела в пространстве.
- 5. Оливо-спинномозговой путь обеспечивает безусловнорефлекторное поддержание тонуса мышц шеи и двигательные акты, направленные па сохранение равновесия.
Оливо-спинномозговой путь начинается от нейронов нижнего оливного ядра продолговатого мозга. Являясь филогенетически новым образованием, нижнее оливное ядро имеет непосредственные связи с корой полушарий лобной доли (корково-оливный путь), с красным ядром (красноядерно-оливный путь) и с корой полушарий мозжечка (оливо-мозжечковый путь). Аксоны клеток нижнего оливного ядра собираются в пучок - оливо-спинномозговой путь, который проходит в переднемедиальном отделе бокового канатика. Он прослеживается только на уровне шести верхних шейных сегментов спинного мозга.
Волокна оливо-спинномозгового тракта посегментно заканчиваются на мотонейронах двигательных ядер передних рогов спинного мозга, аксоны которых в составе передних корешков спинномозговых нервов покидают спинной мозг и направляются к мышцам шеи.
6. Медиальный продольный пучок осуществляет согласованные движения глазных яблок и головы. Эта функция необходима для поддержания равновесия тела. Выполнение данной функции становится возможным только в результате морфофункциональной связи между нервными центрами, обеспечивающими иннервацию мышц глазного яблока (двигательные ядра III, IV и VI пар черепных нервов), центрами, отвечающими за иннервацию мышц шеи (двигательное ядро XI пары и двигательные ядра передних рогов шейных сегментов спинного мозга), центром равновесия (ядро Дей- терса). Координируют работу названных центров нейроны крупных ядер ретикулярной формации - интерстициального ядра (ядра Кахаля) и ядра задней спайки (ядра Дарк- шевича).
Интерстициальное ядро и ядро задней спайки располагаются в ростральном отделе среднего мозга в его центральном сером веществе. Аксоны нейронов этих ядер формируют медиальный продольный пучок, который проходит под центральным серым веществом вблизи срединной линии. Не меняя своего положения, он продолжается в дорсальной части моста и в вентральном направлении отклоняется в продолговатом мозге. В спинном мозге он располагается в переднем канатике, в углу между медиальной поверхностью переднего рога и передней белой спайкой. Прослеживается медиальный продольный пучок только на уровне верхних шести шейных сегментов.
От медиального продольного пучка направляются волокна к двигательному ядру глазодвигательного нерва, иннервирующему большинство мышц глазного яблока. Далее в пределах среднего мозга из состава медиального продольного пучка направляются волокна к нейронам двигательного ядра блокового нерва стороны. Это ядро отвечает за иннервацию верхней косой мышцы глазного яблока.
В мосту в состав медиального продольного пучка вступают аксоны клеток ядра Дейтерса (VIII пара), которые идут в восходящем направлении к нейронам интерстициального ядра. От медиального продольного пучка отходят волокна к нейронам двигательного ядра отводящего нерва (VI пара), отвечающего за иннервацию латеральной прямой мышцы глазного яблока. И, наконец, в пределах продолговатого и спинного мозга от медиального продольного пучка волокна направляются к нейронам двигательного ядра добавочного нерва (XI пара) и двигательным ядрам передних рогов шести верхних шейных сегментов, отвечающих за функцию мышц шеи.
Кроме общей координации работы мышц глазного яблока и головы, медиальный продольный пучок выполняет важную интегративную роль в деятельности мышц глаза. Осуществляя связь с клетками ядра глазодвигательного и отводящего нервов, он обеспечивает согласованную функцию наружной и внутренней прямых мышц глаза, проявляющуюся в сочетанном повороте глаз в сторону. При этом происходит одновременное сокращение латеральной прямой мышцы одного глаза и медиальной прямой мышцы другого глаза.
При поражении интерстициального ядра или медиального продольного пучка происходит нарушение координированной работы мышц глазного яблока. Чаще всего это проявляется в виде нистагма (частые сокращения мышц глазного яблока, направленные в сторону движения, при остановке взгляда). Нистагм может быть горизонтальным, вертикальным и даже ротаторным (вращательным). Нередко указанные нарушения дополняются вестибулярными расстройствами (головокружение) и вегетативными расстройствами (тошнота, рвота и т.д.).
7. Задний продольный пучок осуществляет связи между вегетативными центрами ствола головного и спинного мозга.
Задний продольный пучок (пучок Шютце) берет начало от клеток задних ядер гипоталамуса. Аксоны этих клеток объединяются в пучок лишь на границе промежуточного и среднего мозга. Далее он проходит в непосредственной близости от водопровода среднего мозга. Уже в среднем мозге часть волокон заднего продольного пучка направляется к добавочным ядрам глазодвигательного нерва. В области моста от заднего продольного пучка отходят волокна к слезному и верхнему слюноотделительному ядрам лицевого нерва. В продолговатом мозге ответвляются волокна к нижнему слюноотделительному ядру языкоглоточного нерва и дорсальному ядру блуждающего нерва.
В спинном мозге задний продольный пучок располагается в виде узкой ленты в боковом канатике, рядом с латеральным корково-спинномозговым трактом. Волокна пучка Шютце посегментно заканчиваются на нейронах промежуточно-латеральных промежуточных ядер, являющихся вегетативными симпатическими центрами спинного мозга.
Лишь небольшая часть волокон заднего продольного пучка обособляется на уровне поясничных сегментов и располагается вблизи центрального канала. Этот пучок носит название околоэпендимального. Волокна данного пучка заканчиваются на нейронах крестцовых парасимпатических ядер.
Аксоны клеток парасимпатических и симпатических ядер покидают ствол головного или спинного мозга в составе черепных или спинномозговых нервов и направляются к внутренним органам, сосудам и железам. Таким образом, задний продольный пучок играет очень важную интегративную роль в регуляции жизненно важных функций организма.
Пирамидная и экстрапирамидная системы головного мозга
Экстрапирамидная система это совокупность структур (образований) головного мозга, участвующих в управлении движениями, поддержании мышечного тонуса и позы, минуя кортикоспинальную (пирамидную) систему. Структура расположена в больших полушариях и стволе головного мозга.
Экстрапирамидные проводящие пути образованы нисходящими проекционными нервными волокнами, по происхождению неотносящимися к гигантским пирамидным клеткам (клеткам Беца) коры больших полушарий мозга. Эти нервные волокна обеспечивают связи мотонейронов подкорковых структур (мозжечок, базальные ядра, ствол мозга) головного мозга со всеми отделами нервной системы, расположенными дистальнее.
Экстрапирамидная система проходит через следующие структуры головного мозга: базальные ганглии, красное ядро, четверохолмие, чёрную субстанцию, ретикулярную формацию ствола головного мозга. С ней тесно связаны ядра вестибулярного комплекса, мозжечок, премоторная область коры. Экстрапирамидная система - эволюционно более древняя система моторного контроля по сравнению с пирамидной системой. Имеет особое значение в построении и контроле движений, не требующих активации внимания. Является функционально более простым регулятором по сравнению с регуляторами пирамидной системы.
Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций (смех, плач). Обеспечивает плавность движений, устанавливает исходную позу для их выполнения.
При поражении экстрапирамидной системы нарушаются двигательные функции (например, могут возникнуть гиперкинезы, паркинсонизм), снижается мышечный тонус.
Функционально экстрапирамидная система неотделима от пирамидной системы. Она обеспечивает упорядоченный ход произвольных движений, регулируемых пирамидной системой; регулирует врожденные и приобретенные автоматические двигательные акты, обеспечивает установку мышечного тонуса и поддержание равновесия тела; регулирует сопутствующие движения (например, движения рук при ходьбе) и выразительные движения (мимика).
Пирамидная система, это система нервных структур, поддерживающая сложную и тонкую координацию движений. Пирамидная система это одно из поздних приобретений эволюции. Низшие позвоночные этой системы не имеют, она появляется только у млекопитающих, и достигает наибольшего развития у обезьян и особенно у человека. Пирамидная система играет особую роль в прямохождении. Начинается в коре больших полушарий, на пирамидных клетках (Беца), иннервирует мелкие мышцы, отвечающие за тонкие дифференцированные движения кисти, мимику и речевой акт. Значительно меньшее их количество иннервирует мышцы туловища и нижних конечностей, организуя произвольные движения.
16. Области коры БП. В коре головного мозга вылеляют три проекционные зоны:
Первичная проекционная зона занимает центральную часть ядра мозгового анализатора. Это совокупность наиболее дифференцированных нейронов, в которых происходит высший анализ и синтез информации, возникают четкие и сложные ощущения. К этим нейронам подходят импульсы по специфическому пути передачи импульсов в кору головного мозга (спиноталамический путь).
Вторичная расположена вокруг первичной, входит в состав мозгового отдела анализатора. Обеспечивает сложное восприятие. При поражении этой зоны возникает сложное нарушении функции. Чувствительность к раздражителям при этом обычно не нарушается, но нарушается способность к интерпретации значения раздражителя.
Третичная проекционная зона - ассоциативная - это полимодальные нейроны, разбросанные по всей коре головного мозга. К ним поступают импульсы от ассоциативных ядер таламуса и конвергируют импульсы различной модальности. Обеспечивает связь между различными анализаторами и участвует в формировании условных рефлексов.
Пирамидная и экстрапирамидная системы регуляции движений.
К системам, осуществляющим регуляцию движений, относятся пирамидная и экстрапирамидная системы, а также структуры, ответственные за регуляцию координации движений. Аксоны всех нисходящих путей заканчиваются исключительно на мотонейронах спинного мозга.
• Пирамидная система.
+ Нисходящие пути.
- Корково-спинномозговой путь передний (пирамидный путь передний, tractus corticospinalis ventralis) образован аксонами нейронов, расположенных в двигательной зоне коры [предцентральная извилина (gyrus precentralis)]. Волокна пути проходят через внутреннюю капсулу и в переднем канатике, заканчиваются в передних рогах, посегментно перекрещиваясь.
- Корково-спинномозговой путь латеральный (пирамидный путь латеральный, tractus corticospinalis lateralis) начинается в коре предцентральной извилины, проходит через внутреннюю капсулу и после перекреста в продолговатом мозге проходит в боковом канатике, заканчиваясь в передних рогах.
- Уровни (топика, от гр. topos — место) наиболее частого поражения нервных структур пирамидной системы, регулирующей функцию поперечнополосатых мышц и произвольные движения:
- тела нейронов центрального двигательного анализатора, расположенные преимущественно в предцентральной извилине двигательной зоны коры — пирамидные нейроны;
- корково-ядерные и корково-спинномозговые пути;
- вставочные клетки спинного мозга, через которые пирамидные нейроны влияют на мотонейроны передних рогов спинного мозга и черепных нервов;
- тела мотонейронов передних рогов спинного мозга и двигательных ядер черепных нервов, аксоны мотонейронов, нервно-мышечные синапсы.
• Экстрапирамидная система.
+ Нисходящие пути.
- Красноядерно-спинномозговой путь (пучок фон Монакова, tractus rubrospinalis) — нисходящий проекционный путь экстрапирамидной системы, начинается от красного ядра, проходит в мозговом стволе и боковом канатике и заканчивается в передних рогах.
- Ретикулоспинальный путь (tractus reticulospinalis) — эфферентный путь экстрапирамидной системы; начинается в ретикулярной формации продолговатого мозга, заканчивается в передних рогах спинного мозга. Контролирует тонус скелетной мускулатуры и висцеральные двигательные функции (например, автоматизм дыхания).
+ Уровни (топика) поражения нервных структур экстрапирамидной системы, регулирующих мускулатуру, обеспечивающую непроизвольные («автоматические») движения:
+ нейроны коры (премоторная зона, поясная извилина и др.), подкорковых ядер стриопаллидарной системы, мозжечка, спинного мозга;
+ проводящие пути стриопаллидарной системы (например, красноядерно-спинномозговой или ретикулоспинальный);
• Система координации движений.
Уровни (топика) поражения нервных структур, обеспечивающих координацию движений:
- тела нейронов лобной и височной областей коры головного мозга, ядер мозжечка;
- проводящие пути (от нейронов коры мозга и мозжечка к нейронам гипоталамуса, красного ядра среднего мозга, вестибулярных ядер, ретикулярной формации ствола мозга и др.);
- синаптические структуры.
Управление движениями по мере эволюционной цефализации функций переходило от старых к более новым развивающимся отделам нервной системы. Если рыбы с удаленным головным мозгом свободно плавают, почти не отличаясь в своих движениях от интактных, то уже у амфибий и рептилий структуры заднего и особенно среднего мозга становятся участниками высшей регуляции поведения; у птиц необходимым для двигательной активности оказывается передний мозг, у млекопитающих его большие полушария — кора и подкорковые структуры — осуществляют управление всеми моторными функциями. В этом управлении различают две совместно действующие системы — пирамидную и экстрапирамидную.
Пирамидная система, филогенетически более молодая, возникла с появлением новой коры, в которой она представлена прежде всего гигантскими нейронами пирамидной формы, посылающими аксоны через все отделы головного мозга и по нисходящим путям спинного мозга прямо к его мотонейронам. Эти клетки открыл киевский анатом В. А. Бец (1874). Гигантские пирамидные клетки Беца находятся в V слое сенсомоторной области коры больших полушарий головного мозга. Кроме них в пирамидный тракт посылают свои аксоны менее крупные пирамидные клетки других слоев и зон коры.
Волокна пирамидного тракта проходят через внутреннюю капсулу, ножки мозга и выходят на вентральную поверхность продолговатого мозга. Здесь большинство (70 — 90 %) волокон пирамидного тракта перекрещивается и спускается в спинной мозг как нисходящие пирамидные пути боковых столбов его белого вещества. Неперекрещенные волокна опускаются в спинной мозг по нисходящим пирамидным путям передних столбов своей стороны, но перед тем, как закончиться, они также переходят на противоположную сторону. Таким образом, все пирамидные пути от клеток моторной зоны правого полушария приводят в действие мотонейроны передних рогов левой половины спинного мозга, а идущие от левого полушария — мотонейроны правой половины. Поэтому раздражение моторной зоны одного из полушарий вызывает движения на противоположной стороне тела.
По волокнам пирамидного тракта постоянно протекают потоки импульсов (Э. Эдриан, Д. Моруцци, 1939). Предполагают, что по пирамидному тракту осуществляются два механизма управления движениями: быстро действующий по толстым волокнам (более 10 мкм) для выполнения срочных реакций и медленно действующий по тонким волокнам (менее 5 мкм) для регулирования функционального состояни я спинномозговых центров и перестроек позных рефлексов. Электрофизиологическое исследование передачи возбуждения от пирамидных путей к мотонейронам спинного мозга показало, что она происходит через нейроны его базилярных ядер, Кора задает по пирамидным путям различные двигательные программы ходьбы, прыжков, лазания и т. д. У человека по пирамидным путям выполняются волевые движения под контролем сознания.
Экстрапирамидная система, филогенетически более древняя, в отличие от пирамидной, прямо связанной с исполнительными элементами спинного мозга, соединена с ними через комплекс структур больших полушарий и мозгового ствола. В эту систему входят в основном базальные ганглии, образующие стриопаллидарную группу полосатого тела corpus striatum, включающего хвостатое ядро nucleus caudatum и скорлупу putamen, бледный шар globus pallidum, субталамическое Люисово тело промежуточного мозга, черную субстанцию и отчасти красное ядро среднего мозга. Они представляют собой последовательно возникшие в эволюции супраспинальные уровни координации движений и получают особое развитие у приматов в связи с совершенствованием манипуляторной деятельности передних конечностей, достигшей чрезвычайной тонкости и многогранности у человека. Функции структур, образующих зкстрапирамидную систему, и их взаимодействие отражают эволюционное усложнение управления моторикой. Например, бледный шар у рыб осуществляет регуляцию ритмических движений плавников. У амфибий эта функция бледного шара теряет свое значение и развивающиеся новые структуры тормозят его активность.
На нижних уровнях экстрапирамидной системы красное ядро сдерживает активность разгибателей, активируемых ядром Дейтерса, и уравновешивает их тонус напряжением сгибателей, изменяя их отношения при локомоциях. Выключение связей между ними ведет к описанным выше явлениям децеребра-ционной ригидности, т. е. контр актильно го тонуса. Выключение связей красного ядра с вышележащими структурами ведет к явлениям так называемой восковидной сгибаемости, т. е. пластического тонуса. При этом напряжение сгибателей и разгибателей автоматически уравновешивается и фиксирует конечности и тело в любом придаваемом ему положении.
Из оперированного таким образом животного можно «лепить» какие угодно фигуры, ставить его в самые различные позы, которые оно будет сохранять. Подобное состояние можно наблюдать у человека при некоторых поражениях базальных ганглиев, каталепсии, гипнотическом состоянии, отравлении бульбокапнином. Черная субстанция, бледный шар и полосатое тело образуют все более высокие уровни организации сложных локомоторных актов, устанавливают связи с та л ами чески ми ядра ми и гипоталамусом и получают информацию о внешних событиях и внутреннем состоянии организма, благодаря чему формируют специфические формы вспомогательных движений мимики, жестикуляций и других проявлений эмоций, характерных для инстинктивного поведения.
Совместная деятельность пирамидной и экстрапирамидной систем человека формирует основные уровни построения его движений. Первый наиболее примитивный, руброспиналъный уровень проприоцептивных рефлексов, протекающих с участием красного ядра и спинного мозга без вмешательства сознания, обеспечивает автоматические движения, например ходьбу. Второй более высокий, таламо-паллидарный уровень формирует синергии осознаваемых движений, координированных в результате взаимодействия таламуса и бледного ядра, например выполнение гимнастических упражнений. Третий — пирамидно-стриарный уровень организации движений, сознательно выполняемых с участием моторной коры и полосатого тела под контролем зрения, например рисование картины с натуры. Четвертый — темеиио-премоторный уровень выполнения целенаправленных движений с использованием корковых механизмов выработанных навыков, например забивание гвоздя молотком. Наконец, пятый — высший кортикальный уровень речи и письма — осуществляется второй сигнальной системой мозга человека, например запись протокола проведенного эксперимента или решения математической задачи.
Стратегия поведения определяется текущей мотивацией и генетической программой, выбор тактики зависит от реальных условий обстановки, в соответствии с которыми формируется программа действий. Сенсорный фон действий создается восприятием окружающей среды и отношением к ней. Непрерывные коррекции движений по ходу их выполнения обеспечиваются обратными связями на всех уровнях нервного управления.
38. Базальные ядра больших полушарий (nuclei basalis), подкорковые ядра, базальные ганглии, скопления серого вещества в толще белого вещества больших полушарий головного мозга позвоночных, участвующие в координации двигат. активности и формирования эмоц. реакций. Б. я. вместе с корой мозга составляют клеточное вещество конечного мозга. Состоят из хвостатого ядра, скорлупы (объединяются в полосатое тело), бледного шара (объединяется со скорлупой в чечевицеобразное ядро), ограды, миндалевидного тела. У рыб и земноводных Б. я. представлены только бледным шаром. У пресмыкающихся впервые появляются хвостатое ядро и скорлупа, особенно хорошо развитые у птиц. Б. я. характеризуются множественными афферентными и эфферентными связями с корой больших полушарий, средним и промежуточным мозгом, лимбич. системой и мозжечком. Для нормального функционирования Б. я. исключительно важное значение имеет дофамин, выполняющий роль тормозного медиатора, а также ацетилхолин. У низших позвоночных со слабо развитой корой больших полушарий Б. я. выполняют функцию осн. интегративного аппарата головного мозга, у высших — сохраняют важную роль в регуляции произвольных, движений.
10.1. Общий план строения конечного мозга
Конечный мозг представлен двумя большими полушариями. Большие полушария — самая большая часть головного мозга человека. В норме полушария относительно симметричны и соединены между собой массивным пучком аксонов (мозолистым телом), по которому происходит передача информации из одного полушария в другое. В состав каждого полушария входят базальные ядра, желудочек, белое вешество и плащ, образованный корой.
В соответствии с филогенезом выделяют обонятельный мозг, базальные ядра и кору полушарий (рис. 10.1)
Читайте также: