Фосфолипиды как вторичные посредники. Система кальций-кальмодулин как вторичный посредник
Добавил пользователь Евгений Кузнецов Обновлено: 02.02.2026
1. ЦИКЛИЧЕСКИЕ НУКЛЕОТИДЫ В РОЛИ ВТОРИЧНЫХ ПОСРЕДНИКОВ
цАМФ открыт при изучении стимуляции гликогенолиза
в печени адреналином.
Циклический аденозинмонофосфат (цАМФ) - первое
соединение, которое Сазерленд назвал вторичным
посредником.
Фермент, который образует цАМФ, - аденилатциклаза
Фермент, который разрушает
цАМФ (переводит в
нециклическую форму) -
фосфодиэстераза
Внутриклеточная концентрация цАМФ определяется
скоростью синтеза и распада этого соединения
5. ХАРАКТЕРИСТИКИ цАМФ КАК ВТОРИЧНОГО ПОСРЕДНИКА
В ответ на лиганд
возрастает
концентрация цАМФ
Лиганд должен
стимулировать
аденилатциклазу
в препаратах
чувствительной
ткани
Воспроизведение
эффекта цАМФ с
помощью
проникающего в
клетку аналога
Ингибиторы
фосфодиэстеразы
усиливают эффект
гормона
Повышение
концентрации
цАМФ в ткани
предшествует
биологическому
эффекту
6. ПРИМЕРЫ цАМФ-ЗАВИСИМЫХ ПРОЦЕССОВ
СЕКРЕЦИЯ ЭКЗО- И
ЭНДОКРИННЫХ
ЖЕЛЕЗ
ИЗМЕНЕНИЕ
ПРОНИЦАЕМОСТИ
КЛЕТОЧНЫХ МЕМБРАН
СИНТЕЗ
СТЕРОИДНЫХ
ГОРМОНОВ
цАМФзависимые
процессы
ТРАНСКРИПЦИЯ
ГЕНОВ
ПОДВИЖНОСТЬ,
АГРЕГАЦИЯ КЛЕТОК
ПЕРЕМЕЩЕНИЯ
ВНУТРИКЛЕТОЧНЫХ
СТРУКТУР
9. ПРОТЕИНКИНАЗА А
Протеинкиназа А в
неактивном состоянии
состоит из 2 регуляторных и
2 каталитических
субъединиц.
После присоединения 4
молекул цАМФ к
регуляторным субъединицам
происходит диссоциация
комплекса, и каталитические
субъединицы приобретают
активность
10. БЕЛКИ-СУБСТРАТЫ ДЛЯ ПРОТЕИНКИНАЗЫ А
•БЕЛКИ ИОННЫХ КАНАЛОВ
•ИОНООБМЕННИКИ
•ТРАНСПОРТНЫЕ АТФазы
•БЕЛКИ ЦИТОСКЕЛЕТА
•ФЕРМЕНТЫ, РЕГУЛИРУЮЩМЕ
МЕТАБОЛИЧЕСКИЕ
ПРОЦЕССЫ
•БЕЛКИ РИБОСОМ
•ЯДЕРНЫЕ БЕЛКИ
11. цГМФ
12. ОБРАЗОВАНИЕ И РАСПАД цГМФ
цГМФзависимые
процессы
цГМФ играет важную роль в регуляции Са2+-гомеостаза в
различных типах клеток.
Повышение концентрации цГМФ приводит к понижению
концентрации Са2+ в результате
активации Са2+-АТФ-аз
активации Na/Ca-обменника
подавления рецепторзависимого поступления Са2+ в
клетку
Эти эффекты опосредованы действием протеинкиназы G
на мембранные белки, участвующие в обмене Са2+.
ПК G фосфорилирует Na,K-АТФазу
эпителиоцитов почечных канальцев, что
приводит к торможению насоса.
Результат: ослабление реабсорбции ионов
натрия, а значит и воды из первичной мочи.
15. ПУТИ ОБРАЗОВАНИЯ цГМФ
16. ДВЕ ФОРМЫ ГУАНИЛАТЦИКЛАЗЫ
Мембранная
гуанилатциклаза
Растворимая
форма
гуанилатциклазы
активируется
оксидом азота
ПРОТЕИНКИНАЗА G
17. МЕХАНИЗМ ДЕЙСТВИЯ НАТРИЙУРЕТИЧЕСКОГО ПЕПТИДА
ОСОБЕННОСТИ цГМФ-ЗАВИСИМОЙ
РЕГУЛЯЦИИ
ОТСУТСТВУЮТ G-БЕЛКИ
СВЯЗЫВАНИЕ АГОНИСТА С РЕЦЕПТОРОМ
ВСЕГДА ПРИВОДИТ К АКТИВАЦИИ
ГУАНИЛАТЦИКЛАЗЫ
19. РАСТВОРИМАЯ ФОРМА ГЦ
20. ПРОТЕИНКИНАЗА G
Протеинкиназа G в
неактивной форме
состоит из 2 субъединиц,
имеет 2 регуляторных
центра, с которыми
соединяются 2 молекулы
цГМФ. Кроме того, в ходе
активации этого
фермента не происходит
диссоциация на
субъединицы
21. РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
Факторы роста (GF)
или митогены
стимулируют рост
клетки и ее
прохождение через
митоз.
GF - полипептиды,
состоящие из 50-100
аминокислот.
РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА
1
2
3
1-
внеклеточный N-концевой участок, связывающий
агонисты;
2 - трансмембранный участок;
3 - внутриклеточный тирозинкиназный домен
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ
РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ
(РТК)
ДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ
ТРАНСАВТОФОСФОРИЛИРОВАНИЕ
ТИРОЗИНОВЫХ ОСТАТКОВ
ФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ
СВЯЗЫВАНИЯ БЕЛКОВ-СУБСТРАТОВ,
СОДЕРЖАЩИХ SH2 и SH3 ДОМЕНЫ
ФОСФОРИЛИРОВАНИЕ
СВЯЗЫВАЮЩИХСЯ С РТК БЕЛКОВ
Ключевой компонент сигнального
пути, связанного с рецепторной
тирозинкиназой - Ras-белок.
Ras-БЕЛОК - МОНОМЕРНАЯ ГТФаза
В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ
С ГТФ, В НЕАКТИВНОМ - С ГДФ
АКТИВАЦИЯ И ИНАКТИВАЦИЯ Rаs-БЕЛКА
ОСУЩЕСТВЛЯЕТСЯ ДВУМЯ СПЕЦИФИЧЕСКИМИ
БЕЛКАМИ (GEF (Sos) и GAP)
ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС
ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ
(КИНАЗНЫЙ КАСКАД)
ЭФФЕКТ - ДЕЛЕНИЕ КЛЕТОК
Переход Ras белка из неактивной формы в
активную происходит с участием двух белков.
1- Гуанин-нуклеотид
обменивающий фактор GEF
(Sos) вызывает диссоциацию
GDP от Ras.
2- GTP спонтанно
связывается, а GEF
диссоциирует, освобождая
активный комплекс Ras-GTP.
3 - Гидролиз GTP в сотни раз
усиливается активирующим
GTP-азу белком GAP
Активированный RAS-белок
активирует ПКазу Raf
ПК Raf активирует ПКазу МЕК
(киназа МАР-киназ)
ПК МЕК активирует ПКазу МАР
(митогенактивируемая
протеинкиназа)
ПКаза фосфорилирует множество
различных субстратов, включая
факторы транскрипции
30. РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ
Эти рецепторы при активации
связываются с цитоплазматическими
тирозинкиназами и
образуют сигнальный комплекс.
Рецепторы цитокинов
Рецепторы антигенов на Т- и Влимфоцитах
Fc - рецепторы
32. КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
Известно более 30 БАВ (гормоны, факторы
роста, простагландины, цитокины), действие
которых опосредовано ионами кальция.
Большинство агонистов вызывает повышение
внутриклеточной концентрации ионов кальция
(например, ангиотензин II, эндотелин I,
тромбоксан, брадикинин, гистамин, серотонин и
др.)
Гораздо меньше агентов, приводящих к
снижению внутриклеточной концентрации
ионов кальция (например, простациклин,
простагландин Е1, Na-уретический пептид)
35. Роль ионов кальция как вторичного посредника
•Низкая внутриклеточная концентрация ионов кальция
( 10-7 М)
•Возможность быстро повысить внутриклеточную
концентрацию (до 0,6 - 2 мкМ) ионов кальция
36. Характеристики Ca2+как вторичного посредника
1. В ответ на действие БАВ активируются структуры,
которые поставляют ионы кальция в клетку.
2. В ответ на БАВ увеличивается [Ca2+]in.
3. Снижение активности Ca2+-АТФазы приводит к усилению
эффекта ионов Ca2+.
4. Эффект БАВ воспроизводится с помощью Ca2+-ионофора.
5. Повышение [Ca2+]in предшествует биологическому
эффекту.
37. Кальций-связывающие белки
I.
Кальций-связывающие белки с ферментативной
активностью
II.
Кальций-фосфолипид-связывающие белки
III. Кальций-запасающие белки
IV. Кальций-связывающие белки в составе ионных
каналов и др. ион-транспортных систем
V.
Регуляторные кальций-связывающие белки, не
проявляющие ферментативной активности
I.Кальций-связывающие белки с
ферментативной активностью
кальпаин
Мультисубстратная протеаза
кальцинейрин
Протеинфосфатаза
КАЛЬПАИН
Гетеродимер: большая
каталитическая и малая
регуляторная
субъединицы.
гидролизует белки цитоскелета,
ядерные белки, мембранные
рецепторы, многие ферменты,
транспортные белки
КАЛЬЦИНЕЙРИН
Гетеродимер, состоит из
регуляторной и
каталитической субъединиц,
которые, в свою очередь,
имеют несколько доменов.
Широко представлена в
мозге
Дефосфорилирует различные
функционально важные фосфобелки
по серину и треонину
41. II.Кальций-фосфолипид-связывающие белки
АННЕКСИНЫ (известно более 10)
Содержат 4 Са2+-связывающих домена
В ПРИСУТСТВИИ ИОНОВ КАЛЬЦИЯ СВЯЗЫВАЮТ
ФОСФОЛИПИДЫ МЕМБРАН, ТЕМ САМЫМ
УЧАСТВУЮТ В ПРОЦЕССАХ СЛИЯНИЯ И АГРЕГАЦИИ
МЕМБРАН
ИНГИБИРУЮТ ФОСФОЛИПАЗУ А2
КОНТРОЛИРУЮТ ПРОЛИФЕРАЦИЮ И ДИФФЕРЕНЦИРОВКУ
КЛЕТОК
Во время апоптоза клетки высвобождают
фосфатидилсерин на клеточной поверхности.
Аннексин V, являющийся фосфолипид-связывающим
протеином, в присутствии ионов кальция селективно,
с высокой аффинностью, связывает фосфатидилсерин.
43. III.Кальций-запасающие белки
СИСТЕМА ПАССИВНОГО ЗАБУФЕРИВАНИЯ ИОНОВ КАЛЬЦИЯ (примеры)
КАЛЬСЕКВЕСТРИН
В поперечно-полосатых
мышцах
КАЛЬРЕТИКУЛИН
В СПР
44. IV.Кальций-связывающие белки в составе ион-транспортных систем
45. V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности
Центральное место среди этих белков занимает
КАЛЬМОДУЛИН
Впервые обнаружен Ченгом в мозге КРС в
качестве Са2+-зависимого активатора
фосфодиэстеразы циклических
нуклеотидов.
Впоследствии обнаружен во всех
эукариотических клетках.
Является “E - F hand”протеином.
В структуре белка было
обнаружено 6 -спиралей
(A,B,C,D,E,F). В петлях
полипептидной цепи между
участками C и D, E и F
находятся функциональные
группы с высоким
сродством к кальцию.
- ион Са2+
Спирали E и F расположены
как вытянутые
указательный и большой
пальцы
Связывающие Са2+
участки содержат остатки
кислых аминокислот и
обогащены атомами
кислорода.
6 - 8 атомов кислорода
образуют ячейку,
соответствующую
размеру иону кальция
(r 0,099нм)
49. Са2+-кальмодулин-зависимые
ферменты
процессы
Киназа легких цепей
миозина (в ГМК)
Са-АТФаза
Аденилатциклаза
ФДЭ
Са-СаМ-зависимая
протеинкиназа
Сокращение ГМК
Клеточная
пролиферация
Диссоциация
микротрубочек
Высвобождение
нейромедиаторов
КОМПЛЕКС КАЛЬЦИЙ-КАЛЬМОДУЛИН
ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ
С ФЕРМЕНТОМ-МИШЕНЬЮ
ЧЕРЕЗ Са2+-КАЛЬМОДУЛИНЗАВИСИМУЮ
ПРОТЕИНКИНАЗУ
51. ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ КОМПЛЕКСА Са2+ - КАЛЬМОДУЛИН С ФЕРМЕНТОМ-МИШЕНЬЮ
53. СИГНАЛЬНАЯ СИСТЕМА, ОПОСРЕДОВАННАЯ ИОНАМИ КАЛЬЦИЯ И ПРОДУКТАМИ ГИДРОЛИЗА МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ
ВИДЫ ФОСФОЛИПАЗ
PLА1 - гидролиз эфирной
связи в положении 1
(отщепляет SN-1 ацильную
цепь).
PLА2 - гидролиз эфирной
PLС - гидролиз эфирной связи в положении
3 (гидролизует связь между глицериновым
остатком фосфолипида и полярной фосфатной
группой)
PLD - гидролизует связь между фосфатной
группой и спиртовой группой
связи в положении 2
(отщепляет SN-2 ацильную
цепь, образование свободных
жирных кислот)
МЕТАБОЛИЗМ МЕМБРАННЫХ
ФОСФОИНОЗИТИДОВ
Фосфатидилинозитол(PI)
PI-4-фосфат (PIP)
PI-4,5-бифосфат (PIP2)
57. ПРОДУКТЫ ГИДРОЛИЗА ФОСФОИНОЗИТИДОВ: ВТОРИЧНЫЕ ПОСРЕДНИКИ
Инозитолтрифосфат -
гидрофильная молекула, которая
диффундирует в цитозоль и
взаимодействует с рецепторами
ретикулума
Диацилглицерол - гидрофобная
молекула, которая остается в
мембране. Под действием
фосфолипазы А2 дает арахидоновую
кислоту, которая является
предшественником эйкозаноидов.
58. ИЗОФОРМЫ ФОСФОЛИПАЗЫ С
ФОСФОЛИПАЗА С: изоформы и
ФОСФОЛИПАЗА С активируется через
рецепторы, связанные с G-белками
(Gqбелок)
ФОСФОЛИПАЗА С активируется
благодаря фосфорилированию ее
тирозинкиназой.
63. ПРОЦЕССЫ, В КОТОРЫХ УЧАСТВУЕТ ПРОТЕИНКИНАЗА С
Протеинкиназа С
R
состоит из двух доменов -
регуляторного (R) и каталитического
(C)
С
2Zn2+
Каталитический домен: центр,
связывающий АТФ и белок-субстрат.
Са2+
АТФ
Белоксубстрат
Неактивная протеинкиназа С -
цитозольная, при активации
становится мембранносвязанной
ДАГ
Регуляторный домен: мотив цинковые
пальцы и участок с высоким
сродством к Са2+
67. Газовые посредники - новый класс сигнальных молекул
68. Общие свойства газовых посредников
Небольшие липофильные молекулы, свободно
проникают через мембраны
Функционируют без участия мембранных рецепторов
Проявляют эндокринный, паракринный и аутокринный
эффекты
Образуются эндогенно благодаря специальным
ферментам
Имеют специфическую функцию при физиологически
значимых концентрациях.
Функция
газового
посредника
может
быть
имитирована экзогенно его аналогом (применение
доноров)
Клеточные эффекты газовых посредников могут быть
прямыми
или
опосредованными
вторичными
посредниками, но всегда есть специфические
клеточные и молекулярные мишени
72. РОЛЬ ОКСИДА АЗОТА
Регулятор
переноса
кислорода
эритроцитами
Участник
межнейрональной
передачи
информации
Фактор, влияющий на
гипотоническую
устойчивость
эритроцитов
NO
Регулятор
сократимости
скелетных мышц
Ингибирует
агрегацию
тромбоцитов
Участник
иммунных
реакций
Эндогенный
вазодилятатор
NO ВЫРАБАТЫВАЕТСЯ ВО МНОГИХ КЛЕТКАХ, НО
СУЩЕСТВУЮТ ТРИ КАТЕГОРИИ КЛЕТОК, В КОТОРЫХ
ПРОЯВЛЯЮТСЯ ФУНКЦИИ ЭТОЙ МОЛЕКУЛЫ
ЭНДОТЕЛИАЛЬНЫЕ КЛЕТКИ (РАССЛАБЛЕНИЕ ГМК)
НЕЙРОНЫ (ПЕРЕДАЧА СИГНАЛА)
ИММУНОКОМПЕТЕНТНЫЕ КЛЕТКИ (УЧАСТИЕ В
ИММУННОМ ОТВЕТЕ)
74. ИЗОФОРМЫ NO-СИНТАЗЫ
ОКСИД АЗОТА ВЫРАБАТЫВАЕТСЯ ИЗ L-АРГИНИНА ПОД
ДЕЙСТВИЕМ NO-СИНТАЗЫ
Конститутивная форма
•В эндотелиальных клетках
и нейронах
•Активность зависит от Са2+
•Связана с мембраной
•Концентрация образуемого
NO невелика
•Функции: расслабление
ГМК сосудов,
межнейрональная передача
Индуцибельная форма
•В макрофагах
•Активируется цитокинами
•Цитозольный фермент
•Концентрация образуемого
NO большая
•Функции: цитотоксическое и
цитостатическое действие
на атакуемые макрофагами
клетки
Эндотелиальная NO-синтаза
нерастворимый фермент с молекулярной массой
около 135 кДа.
NO, продуцируемый эндотелиальной синтазой,
регулирует тонус кровеносных и
лимфатических сосудов,
предупреждает тромбообразование.
77. РАСТВОРИМАЯ ФОРМА ГЦ
Нейрональная NO-синтаза
растворимый гомодимер с молекулярной массой
около 150 кДа
содержит кальмодулин-связывающий центр,
осуществляющий Са+-зависимую регуляцию
синтеза NO
содержит атом железа, входящий в состав
гемовой простетическкои группы
Функции нейронального NO:
контролирует осцилляторную активность нейронов
является медиатором ноцицепции,
термочувствительности, обоняния
регулирует выход нейромедиаторов
играет центральную роль в процессах обучения и
памяти
Индуцибельная NO-синтаза
Кальций-независимая
индуцибельная NO-синтаза
обнаружена в макрофагах,
гепатоцитах, фибробластах,
миоцитах.
Индуцибельная NO-синтаза
При активации синтеза фермента образование
NO возрастает в десятки раз и максимальных
значений достигает через часы.
iNOS индуцируется в зараженных вирусом
клетках.
iNOS стимулирует образование нестабильных
свободных радикалов
85. Субстрат для образования СО - молекула гема, являющаяся небелковым компонентом гем-протеинов (гемоглобин, миоглобин, каталаза,
86. Продукция СО продемонстрирована в сосудах, нервной ткани, печени, почках, коже, кишечнике, легких, желудочках сердца, эпифизе,
ФУНКЦИИ СО
выступает в роли нейротрансмиттера
влияет на тонус гладкой мускулатуры сосудов
обладает противосвертывающим эффектом
.
играет сигнальную роль в иммунной системе
Пути образования H2S в организме:
Ферментативный - из L-цистеина
цитозольными пиридоксаль-5’-фосфат
(витамин В6)-зависимыми ферментами цистатионин -синтазой (CBS) и цистатионин лиазой (CSE);
Неферментативный - синтез H2S при
окислении глюкозы в присутствии
элементарной серы
90. Метаболизм сероводорода
91. Известные молекулярные мишени H2S
92. ЭФФЕКТЫ СЕРОВОДОРОДА
Регуляция
нейрональной
активности
H2S
Ингибирует
агрегацию
тромбоцитов
Провоспалительный
агент, поскольку
усиливает выработку
провоспалительных
цитокинов
Эндогенный
вазодилятатор
Рис. 6.2. Опосредование гормонального сигнала системой фосфолипаза С-инози- тол-3-фосфат.
Образование гормон-рецепторного комплекса при участии регуляторного G-белка активирует мембранную фосфолипазу С, вызывающую гидролиз фосфолипидов мембраны с образованием двух вторичных посредников: инозитол-3-фосфата и диацил глицерола. Инозитол-З-фосфат ведет к выходу Са 2+ из внутриклеточных депо. Связывание ионизированного кальция со специализированным белком кальмодулином активирует протеинкиназы и вызывает фосфорилирование внутриклеточных структурных белков и ферментов. Диацилглицерол повышает сродство протеинкиназы С к Са 2+ , способствуя ее активации, что также завершается процессами фосфорилирования белков. Диацилглицерол одновременно реализует другой путь опосредования гормонального эффекта, активируя фосфолипазу А-2 и образование простаноидов.
ский эффект гормона. Через рассмотренную систему вторичных посредников реализуются эффекты адреналина (при связи с альфа-адренорецептором), вазопрессина (при связи с V-1-рецептором), ангиотензина-II, соматостатина, окситоцина.
4. Система кальций—кальмодулин. Ионизированный кальций поступает в клетку после образования гормон-рецепторного комплекса либо из внеклеточной среды за счет активирования медленных кальциевых каналов мембраны (как это происходит, например, в миокарде), либо из внутриклеточных депо под влиянием инозитол-3-фосфата. В цитоплазме немышечных клеток кальций связывается со специальным белком-кальмодули- ном, а в мышечных клетках роль кальмодулина выполняет тропонин С. Связанный с кальцием кальмодулин изменяет свою пространственную организацию и активирует многочисленные протеинкиназы, обеспечивающие фосфорилирование, а следовательно изменение структуры и свойств белков. Кроме того комплекс кальций—кальмодулин активирует фосфодиэстеразу цАМФ, что подавляет эффект циклического соединения как вторичного посредника. Вызываемое гормональным стимулом кратковременное увеличение в клетке кальция и его связывание с кальмодулином является пусковым стимулом для многочисленных физиологических процессов — сокращения мышц, секреции гормонов и выделения медиаторов, синтеза ДНК, изменения подвижности клеток, транспорта веществ через мембраны, изменения активности ферментов.
6.1.1.2. Взаимосвязи вторичных посредников
В клетках организма присутствуют или могут образовываться одновременно несколько вторичных посредников. В связи с этим между вторичными посредниками устанавливаются различные взаимоотношения: 1) равнозначное участие, когда разные посредники необходимы для полноценного гормонального эффекта; 2) один из посредников является основным, а другой лишь способствует реализации эффектов первого; 3) посредники действуют последовательно (например, инозитол-3-фосфат обеспечивает освобождение кальция, диацилглицерол облегчает взаимодействие кальция с протеинкиназой С); 4) посредники дублируют друг друга для обеспечения избыточности с целью надежности регуляции; 5) посредники являются антагонистами, т. е. один из них включает реакцию, а другой — тормозит (например, в гладких мышцах сосудов инозитол-3-фосфат и кальций реализуют их сокращение, а цАМФ — расслабление).
6.1.2. Механизм действия стероидных гормонов
Стероидные гормоны (рис. 6.3) обладают двумя путями действия на клетки: 1) классическим геномным или медленным и 2) быстрым негеномным.
6.1.2.1. Геномный механизм действия
Геномный механизм действия на клетки-мишени начинается трансмембранным переносом молекул стероидных гормонов в клетку (благодаря их растворимости в липидном бислое клеточной мембраны), с последующим связыванием гормона с цитоплазменным белком-рецептором. Эта связь с рецепторным белком необходима для поступления стероидного гормона в ядро, где происходит его взаимодействие с ядерным рецептором. Последующее взаимодействие комплекса гормон—ядерный рецептор с хроматиновым акцептором, специфическим кислым белком и ДНК влечет за собой: активацию транскрипции специфических мРНК, синтез транспортных и рибосомных РНК, процессинг первичных РНК-транскриптов и транспорт мРНК в цитоплазму, трансляцию мРНК при достаточном уровне транспортных РНК с синтезом белков и ферментов в рибосомах. Все эти явления требуют длительного (часы, сутки) присутствия гормон-рецеп- торного комплекса в ядре.
6.1.2.2. Негеномный механизм действия
Эффекты стероидных гормонов проявляются не только спустя несколько часов, что требуется для ядерного влияния, часть из них проявляется очень
быстро, в течение нескольких минут. Это такие эффекты, как повышение проницаемости мембран, усиление транспорта глюкозы и аминокислот, освобождение лизосомальных ферментов, сдвиги энергетики митохондрий. К числу быстрых негеномных эффектов стероидных гормонов относятся, например, увеличение в течение 5 мин после введения человеку альдостерона общего периферического сосудистого сопротивления и артериального давления, изменение транспорта натрия через мембрану эритроцитов (вообще лишенных ядра) под влиянием альдостерона в опытах in vitro, быстрый вход Са 2+ в клетки эндометрия под влиянием эстрогенов и др. Механизм негеномного действия стероидных гормонов заключается в связывании на плазматической мембране клетки со специфическими рецепторами и активации каскадных реакций систем вторичных посредников, например фосфолипазы С, инозитол-3-фосфата, ионизированного Са 2+ , протеинкиназы С. Под влиянием стероидных гормонов в клетке может увеличиваться содержание цАМФ и цГМФ. Негеномный эффект стероидных гормонов
1 — классический геномный путь действия (гормон проникает через клеточную мембрану и цитоплазму в ядро, где после взаимодействия с ядерным рецептором воздействует на гены-ми- шени, активируя их). 2а и 2б — негеномные пути действия через мембранные рецепторы: 2а — пути, связанные с мембранным ферментом и образованием вторичного посредника, ведущего к активации протеинкиназ. Последние через фосфорилирование в ядре белка-коактиватора (БКА) активируют гены-мишени; 26 — пути, связанные с ионными каналами клеточной мембраны, в результате чего гормон-рецепторный комплекс активирует ионные каналы, меняя возбудимость клетки. 3 — альтернативный негеномный путь действия (молекула гормона, проникая через мембрану в цитоплазму, взаимодействует с цитозольным рецептором, что приводит к активации цитозольных киназ.
может быть реализован и после их связывания с цитоплазматическими рецепторами. Часть негеномных эффектов стероидных гормонов осуществляется благодаря их взаимодействию с рецепторами, связанными с воротным механизмом ионных каналов мембран нервных клеток, являясь тем самым модуляторами, например, глицин-, серотонин- или гамма-аминобу- тиратергических нейронов. Наконец, растворяясь в липидном бислое мембраны, стероидные гормоны могут менять физические свойства мембраны, такие как ее текучесть или проницаемость для гидрофильных молекул, что также является негеномным эффектом.
Таким образом, механизмы действия гормонов разной химической структуры имеют не только различия, но и общие черты. Как и стероиды, пептидные гормоны обладают способностью избирательно влиять на транскрипцию генов в ядре клетки. Этот эффект пептидных гормонов может быть реализован не только с поверхности клетки при образовании вторичных посредников, но и путем поступления пептидных гормонов внутрь клетки за счет интернализации гормон-рецепторного комплекса.
6.L3. Саморегуляция чувствительности эффектора к гормональному сигналу
Изменение характера и интенсивности регуляторных сигналов, поступающих к клетке, ведет к адаптивной перестройке ее рецепторных структур за счет специальных механизмов саморегуляции, изменяющих чувствительность клетки к этому сигналу. Так, при избыточности внеклеточного гуморального сигнала формируется десенситизация соответствующего рецептора, т. е. уменьшение его чувствительности к гуморальному регулятору из-за ослабления сродства рецептора к молекуле лиганда или числа рецепторных молекул. Механизм десенситизации рецепторов, реализующих регуляторный эффект через системы вторичных посредников, представлен на рис. 6.4. При наличии во внеклеточной среде избыточного гуморального сигнала и/или увеличении времени связывания лиганда с рецептором активируемая лиганд-рецепторным взаимодействием протеинкиназа плазматической мембраны фосфорилирует рецептор, в результате его аффинность к лиганду уменьшается в 2—5 раз, что предотвращает чрезмерность регуляторного эффекта. Если этого снижения чувствительности рецептора недостаточно, рецептор фосфорилируется другой протеинкиназой, активируемой соответствующим вторичным посредником. Следствием такого фосфорилирования является нарушение сопряжения рецепторов с G-белками и нарушение эффектов G-белков (активирующих или ингибирующих) на системы вторичных посредников и ионные каналы. Наконец, в случаях недостаточной эффективности двух указанных механизмов десенситизации гормон-рецепторные комплексы интернализуются внутрь клетки с образованием рецепторосомы (вакуоль с рецептором). В последующем, если уровень гуморального регулятора во внеклеточной микросреде снизится в течение 2—3 ч, интернализированный рецептор может вновь встроиться в мембрану, а если повышенная концентрация регулятора сохраняется — рецепторосомы сливаются с лизосомами и рецепторы разрушаются. Десенситизация рецепторов может быть гомологической, проявляющейся в снижении чувствительности только к тому гуморальному регулятору, избыток которого ее вызвал, и гетерологической, заключающейся в снижении чувствительности и к другим гуморальным факторам регуляции, но также реализующим эффект через связанные с G-белком рецепторы и те же вторичные посредники.
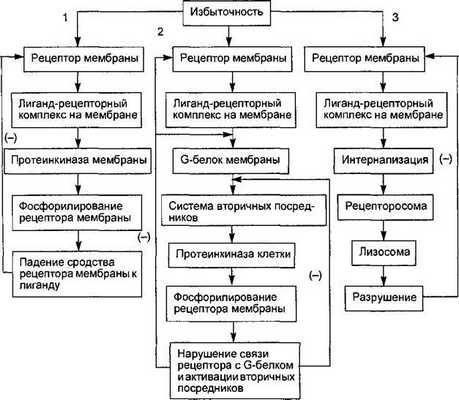
Рис. 6.4. Схема трех путей десенситизации клетки к избыточному внешнему регуляторному сигналу.
1. Снижение сродства рецептора к лиганду из-за фосфорилирования рецептора мембранной протеинкиназой. 2. Нарушение связи молекулы рецептора с G-белком при фосфорилировании рецептора цитоплазматическими протеинкиназами. 3. Интернализация лиганд-рецептор- ного комплекса и разрушение рецептора ферментами лизосом. Стрелками со знаком (—) обозначено подавление процесса взаимодействия.
Чувствительность клеток к внешним регуляторным сигналам зависит также и от числа так называемых свободных, или резервных, рецепторов, т. е. доступных для лиганда. Увеличивая синтез этих мембранных структур или их «всплывание» на поверхность мембраны, клетка способна повышать чувствительность к регуляторному сигналу (сенситизация), например в случаях недостаточного количества молекул гуморального регулятора во внеклеточной микросреде. Повышение чувствительности к регуляторному сигналу является также следствием возросшего сопряжения рецепторов с G-белком и системой вторичных посредников.
Клетки организма, как правило, используют не один, а несколько путей реализации одного гуморального регуляторного сигнала, в связи с чем для каждого лиганда (например, молекул одного гормона) на мембранах существуют несколько типов рецепторов. Так, для ацетилхолина, гистамина, катехоламинов, опиоидных пептидов, серотонина, вазопрессина иденти
фицировано два (вазопрессин), три (опиоиды), четыре (мускариновые для ацетилхолина) и более (катехоламины, серотонин) мембранных рецепторов. Соответственно, меняя за счет саморегуляции чувствительность рецепторов одного типа, клетка изменяет характер функционирования и способна компенсировать неадекватные сдвиги в регуляторных влияниях.
Одним из регуляторных эффектов, реализуемых внутри клетки с помощью систем вторичных посредников мембранного лиганд-рецепторного взаимодействия, является изменение функционального состояния генома клетки и биосинтеза белка. Но и ядро клетки способно менять состояние плазматической мембраны, ее чувствительность к сдвигам внеклеточной среды, в том числе и изменению в ней количества и характера регуляторных сигналов. В клетках под контролем генома происходит синтез специальных белков — инверторов, оказывающих различное влияние на функциональное состояние мембраны в зависимости от адаптивно-компенсаторных потребностей клетки — изменение активности рецепторов, состояния ионных каналов и насосов. Эти белки могут вести к гиперполяризации клеточной мембраны, ослаблению функциональной активности клетки, с одновременным повышением синтеза белка и процессов регенерации внутриклеточных структур.
Понятие о вторичных посредниках. Аденилатциклазная система
Вторичные (внутриклеточные) мессенджеры - малые молек или ионы, внутриклет концентрац кот повышает ответ на соединение рецептора с сигнальной молекулой.
Роль вторичных мессенджеров выполняют молекулы и ионы:
• ц-АМФ (циклический аденозин-3',5'-монофосфат);
• ИФ3 (инозитол-1,4,5-трифосфат);
•ДАГ
•ц-ГМФ
• Са 2 +.
Аденилатциклазная система включает:
- интегральные белки цитоплазматической мембраны:
• Rs - рецептор первичного мессенджера - активатора, рецептор первичного мессенджера - ингибитора АЦС;
• фермент аденилатциклазу (АЦ).
- «заякоренные» белки:
• Gs - ГТФ-связывающие белки
• цитозольный фермент протеинкиназу А (ПКА).
Рис. 4.11. Функционирование аденилатциклазной системы
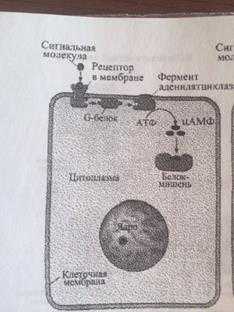
Когда соответствующий лиганд (например, адреналин) взаимодействует с рецептором, происходяi конформационные изменения рецепторного белка. Структурно перестроенный G-белок активизирует аденилатциклазу - фермент, располагающийся в мембране. Активизированная аденилатциклаза катализирует превращение АТФ в ц-АМФ.
В системе ц-АМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са 2+ из эндоплазматической сети. Повышение концентрации Са 2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке.
Участие аденилатциклазной системы в регуляции экспрессии генов. Многие белковые гормоны: глюкагон, вазопрессин, паратгормон и др. могут не только изменять скорости реакций путем фосфорилирования ферментов, но и увеличивать или уменьшать их количество, регулируя экспрессию генов.
Аденилатциклазная система
Циклический АМФ (циклоАМФ, цАМФ) образуется в клетке, когда действуют гормоны гипофиза (ТТГ, ЛГ, МСГ, ФСГ. АКТГ), кальцитонин, соматостатин, глюкагон, паратгормон, адреналин (через α2- и β-адренорецепторы), вазопрессин (через V2-рецепторы). Этапы передачи сигнала:
1) Взаимодействие лиганда с рецептором приводит к изменению конформации последнего.
2)Это изменение передается на G-белок, который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ.В составе G-белков α-субъединицы бывают 2х типов по отношению к аденилатциклазе: активирующие αS и ингибирующие αI.
3)В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно на α-субъединице ГДФ заменяется на ГТФ.
4)Активированная таким образом αS-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ.
Если в действо была вовлечена αI-субъединица - она ингибирует аденилатциклазу, все останавливается.
5) Циклический АМФ - вторичный мессенджер - в свою очередь, взаимодействует с протеинкиназой А и активирует ее. Протеинкиназа А фосфорилирует ряд ферментов, среди которых киназа фосфорилазы гликогена, гликогенсинтаза, ТАГ-липаза.
6)Наработка цАМФ продолжается некоторое время, пока α-субъединица, которая является ГТФ-азой, отщепляет фосфат от ГТФ.
7) Как только ГТФ превратился в ГДФ, то α-субъединица инактивируется, теряет свое влияние на аденилатциклазу, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение.
8)Гормон отрывается от рецептора еще раньше:
- если концентрация гормона в крови велика, то следующая его молекула присоединится к рецептору через малый промежуток времени и повторный запуск АЦ-механизма произойдет быстро - в клетке активируются соответствующие процессы.
- если гормона в крови мало - для клетки наступает некоторая пауза, изменения метаболизма нет. Механизм наработки цАМФ связан с активацией фермента аденилатциклазы и называется аденилатциклазный механизм.
Гормоны, для которых циклический АМФ служит в качестве вторичного медиатора:
Гормон Мишень: орган/ткань Типичное действие
Адренокортикотропный гормон Кора надпочечников Продуцирование кортизола
Кальцитонин Кости Концентрация кальция в сыворотке крови
Катехоламины (b-адренергические) Сердце ЧСС, сократимость миокарда
Хорионический гонадотропин Яичники, семенники Продуцирование половых гормонов
Фолликулостимулирующий гормон Яичники, семенники Гаметогенез
Глюкагон Печень Гликогенолиз, высвобождение глюкозы
Лютеинизирующий гормон Яичники, семенники Продуцирование половых гормонов
Рилизинг-фактор лютеинизирующего гормона Гипофиз Высв-е лютеинизирующего гормона
Меланоцитстимулирующий гормон Кожа (меланоциты) Пигментация
Гормон паращитовидных желез Кости, почки Концентрация Са и Р в сыворотке крови
Простациклин, простагландин Тромбоциты Агрегация тромбоцитов
Тиреотропный гормон Щитовидная железа Продуцирование и высвобождение Тз и Т 4
Рилизинг-фактор тиреотропного гормона Гипофиз Высвоб-е тиреотропного гормона
Вазопрессин Почки Концентрация мочи
По этому механизму, который получил название кальций-фосфолипидный механизм, действуют вазопрессин (через V1-рецепторы), адреналин (через α1-адренорецепторы), ангиотензин II.
Принцип работы этого механизма совпадает с предыдущим, но вместо аденилатциклазы мишеневым ферментом для α-субъединицы служит фосфолипаза С. Фосфолипаза С расщепляет мембранный фосфолипид фосфатидилинозитолдифосфат (ФИФ2) до вторичных мессенджеров инозитолтрифосфата (ИФ3) и диацилглицерола (ДАГ).
Этапы передачи сигнала:
1)Взаимодействие гормона с рецептором приводит к изменению конформации последнего.
2) Это изменение передается на G-белок, который состоит из трех субъединиц (αP, β и γ), α-субъединица связана с ГДФ.
3)В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно на αP-субъединице ГДФ заменяется на ГТФ.
4)Активированная таким образом αP-субъединица стимулирует фосфолипазу С, которая начинает расщепление ФИФ2 до двух вторичных мессенджеров - ИФ3 и ДАГ.
5)Инозитолтрифосфат открывает кальциевые каналы в эндоплазматическом ретикулуме, что вызывает увеличение концентрации ионов Cа2+. Диацилглицерол совместно с ионами Са2+ активирует протеинкиназу С. Кроме этого, диацилглицерол имеет и другую сигнальную функцию: он может распадаться на 1-моноацилглицерол и полиеновую жирную кислоту (обычно арахидоновую), из которой образуются эйкозаноиды.
6)Протеинкиназа С фосфорилирует ряд ферментов и в целом участвует в процессах клеточной пролиферации. Накопление ионов Са2+ в цитоплазме вызывает активацию определенных кальций-связывающих белков (например, кальмодулина).
7)Гидролиз ФИФ2 продолжается некоторое время, пока αP-субъединица, которая является ГТФ-азой, отщепляет фосфат от ГТФ.
8)Как только ГТФ превратился в ГДФ, то αP-субъединица инактивируется, теряет свое влияние на фосфолипазу C, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение.
9) Гормон отрывается от рецептора еще раньше:
- если концентрация гормона в крови велика, то следующая его молекула присоединится к рецептору через малый промежуток времени и повторный запуск механизма произойдет быстро - в клетке активируются соответствующие процессы.
- если гормона в крови мало - для клетки наступает некоторая пауза, изменения метаболизма нет.
Гуанилатциклазный механизм пока изучен недостаточно подробно, но известно, что по нему работает атриопептин (предсердный натрийуретический пептид), оксид азота (NO) и другие, пока малоизученные, соединения.
Гуанилатциклазный механизм передачи сигнала в общих чертах схож с аденилатциклазным: после появления какой-либо сигнальной молекулы сигнал воспринимается ферментом гуанилатциклазой и далее передается при помощи вторичного мессенджера цГМФ. Он воздействует на протеинкиназу G, которая фосфорилирует определенные белки, чем изменяет активность клетки.Но, в отличие от аденилатциклазы, данный фермент имеет четыре разновидности, три из которых связаны с мембраной, четвертый - цитозольный:
мембраносвязанные формы гуанилатциклазы работают как рецепторы, обладающие каталитической активностью.
цитозольный фермент является димером и содержит в своем составе гем, он непосредственно взаимодействует в цитозоле с сигнальной молекулой (например, оксидом азота). Система, генерирующая цГМФ как вторичный посредник, сопряжена с гуанилатциклазой. Этот фермент катализирует реакцию образования цГМФ из ГТФ (подобно аденилатциклазе). Молекулы цГМФ могут активировать транспортные системы мембран клеток или активируют цГМФ-зависимую протеинкиназу G, которая участвует в фосфорилировании других белков в клетке. Циклические нуклеотиды запускают каскады реакций аденилатциклазного или гуанилатциклазного механизмов регуляции активности ферментов. Одна молекула гормона, активирующая рецептор, может «включать» несколько G-белков. Каждый из них в свою очередь активирует несколько молекул аденилатциклазы с образованием тысяч молекул цАМФ или цГМФ. Образующийся вторичный посредник усиливает сигнал в тысячу раз. Суммарное усиление сигнала равно 106 - 107 раз. Снятие гормонального сигнала достигается уменьшением концентрации вторичного посредника. Реакции превращения цАМФ или цГМФ в неактивные метаболиты АМФ или ГМФ катализируют ферменты фосфодиэстеразы. К настоящему времени получено много данных, что цГМФ принадлежит самостоятельная роль в регуляции функции клеток. В частности, в почках и кишечнике он контролирует ионный транспорт и обмен воды, в сердечной мышце служит сигналом релаксации и т.д. Гуанилатциклаза открыта во многих органах (сердце, легкие, почки, надпочечники, эндотелий кишечника, сетчатка и др.), что свидетельствует о широком ее участии в регуляции внутриклеточного метаболизма, опосредованном через цГМФ. Мембраносвязанный фермент активируется через соответствующие рецепторы короткими внеклеточными пептидами в частности гормоном предсердным натрийуретическим пептидом.
Инозитолтрифосфат - это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы "С", который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора. Функционирование инозитолфосфатной системы трансмембранной передачи сигнала обеспечивают: R (рецептор), фосфолипаза С, Gplc - белок, активирующий фосфолипазу С, белки и ферменты мембран и цитозоля. Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона (Пример - АКТГ). В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30 % состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс "Са+2-кальмодулин" становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы. В разных клетках при воздействии комплекса "Са+2-кальмодулин" на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Каскадный механизм передачи гормональных сигналов. Концепция первого и второго посредников. Циклические нуклеотиды и кальций как вторичные посредники
Стероиды и другие липофильные гормоны легко проникают внутрь клеток через поверхностную мембрану, а все остальные гормоны или вообще не проходят через липидную фазу мембраны или проходят с трудом. Липофильные гормоны, по-видимому, прямо воздействуют на генетический аппарат клетки. Большинство же гидрофильных гормонов вызывает более быстрые и кратковременные реакции, влияя, как правило, на обмен веществ. Действие гормонов этой группы опосредуется через рецепторные молекулы на поверхности клеток-мишений и прекращается вскоре после их недолговременного взаимодействия. Взаимодействие гормона с рецептором ведет к запуску внутриклеточного каскадного механизма, усиливающего исходный сигнал. Каскадный механизм представляет собой последовательность биохимических реакций приводящих к образованию в цитозоле большого числа молекул внутриклеточного регулятора. В этой схеме гормон называют внеклеточным или первым посредником, а молекулу (или комплекс молекул) внутриклеточного регулятора, образующегося после взаимодействия гормона с рецептором, - внутриклеточным или вторым посредником. Таким образом, действие многих (главным образом, гидрофильных) гормонов зависит от образования в клетке-мишени второго (а иногда и третьего) посредника, который определяет реакцию внутриклеточных процессов на гормон. Благодаря каскадному усиливающему эффекту клетки-мишени чрезвычайно чувствительны к своему гормону. Некоторые из них реагируют на концентрацию гормонов в плазме крови равную всего 10 -12 М. Если бы наши вкусовые рецепторы обладали такой же чувствительностью к сахару, мы могли бы обнаруживать его присутствие в кофе или чае после растворения щепотки сахара в объеме равном плавательному бассейну.
Интересно, что при большом числе гормонов и гормональных эффектов для реализации последних требуется, по-видимому, очень мало посредников. У самых различных животных до последнего времени были известны только два совершенно разных типа вторых посредников. Один из них - это ионы кальция, которые как мы уже убедились, регулируют целый ряд физиологических функций. К другому типу вторых посредников относятся циклические монофосфаты нуклеотидов, главным образом циклический аденозинмонофосфат (цАМФ) и очень сходный с ним циклический 3'5'-гуанозинмонофосфат (цГМФ). Недавно была открыта еще одна, более сложная система посредников, в которой участвуют фосфолипиды мембран.
Циклический АМФ синтезируется из АМФ ферментом аденилатциклазой (соответственно цГМФ из ГМФ гуанилатциклазой). При действии гуморальных сигналов концентрация в клетке цАМФ или цГМФ (обычно 10 -6 М) должна а может за несколько секунд увеличиться в 5 раз. Многие гормоны и локальные химические медиаторы действуют на клетку, изменяя концентрацию цАМФ (цГМФ), причем они это делают, активируя (реже ингибируя) аденилатциклазу через рецепторы, сопряженные с G-белками. Таким образом, регуляторный GTP-связывающий белок может активировать либо определенные ионные каналы в случае химической передачи в нервной системе, либо фермент аденилатциклазу и повысить уровень циклических нуклеотидов. Разные клетки-мишени, по-разному реагируют на изменение внутриклеточной концентрации цАМФ (цГМФ) и напротив, разные гормоны, если их взаимодействие со своими рецепторами приводит к активации аденилатциклазы в конкретной клетке, обычно будут вызывать одну и ту же реакцию. Например, в жировых клетках аденилатциклазу активируют по крайней мере четыре разных гормона (адреналин, глюкагон, АКТГ, СТГ), и все они вызывают расщепление триглицеридов (резервная форма жира) до жирных кислот. Таким образом различные рецепторы для этих гормонов активируют общий пул аденилатциклазы. Увеличение внутриклеточного уровня ц-АМФ активирует фермент протеинкиназу А. Этот фермент фосфорилирует неактивные белки ферменты, переводя их тем самым в активное состояние. Расщепляются (гидролизуются до соответствующих монофосфатов) циклические нуклеотиды ферментами цАМФ- и цГМФ-специфичными фосфодиэстеразами. Растительные алкалоиды из листьев чая и зерен кофе кофеин и теофиллин (группа метилксантинов) являются ингибиторами фосфодиэстераз и способны стимулировать обменные процессы, повышая внутриклеточную концентрацию циклических нуклеотидов.
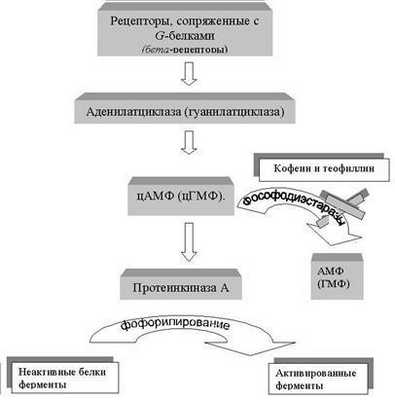
Рис. Каскадный механизм внутриклеточной передачи гормонального сигнала с циклическими монофосфатами в качестве вторых посредников
В последние десятилетия ион Са 2+ привлекает большое внимание как очень важный и вездесущий регулятор клеточных процессов, а также как один из посредников, связывающих внутриклеточные ответы с внешними сигналами, воспринимаемыми плазматической мембраной и ее рецепторами. Известно множество процессов, регулируемых или запускаемых повышением концентрации Са 2+ в клетке: высвобождение нейромедиаторов в синапсах, мышечное сокращение, клеточное деление, различные реакции промежуточного обмена, окислительное фосфорилирование и репликацию ДНК.
Существуют два источника повышения концентрации кальция в цитозоле. Во многих клетках, в том числе мышечных волокнах, ионы кальция освобождаются при стимуляции из внутриклеточных депо, главным образом, из эндоплазматического (саркоплазматического) ретикулума. Второй путь повышения концентрации ионов кальция в цитоплазме - поступление их из внеклеточной среды через кальциевые каналы. Кальциевые каналы могут открываться при деполяризации мембраны (потенциалзависимые) или воздействии химических стимулов. При поступлении в клетку большая часть ионов кальция быстро связывается в цитозоле с анионными участками белковых молекул. Основную массу иммобилизованных (связанных) ионов кальция связывают молекулы, которые просто действуют как буферы, регулирующие концентрацию свободных ионов Са 2+ . Однако некоторые белки служат специфическими рецепторами этих ионов и активируются ими. Таковым, например является кальмодулин. Активированный комплекс Са 2+ -кальмодулин присоединяется к молекулам неактивных или малоактивных ферментов, что приводит к возрастанию активности последних. Известна, например, кальмодулин-зависимая фосфодиэстераза (ФДЭ 1) и ее активность можно заингибировать винпоцетином (кавинтон). Винпоцетин повышает концентрацию циклических нуклеотидов в гладкомышечных клетках сосудов и, благодаря этому, улучшает церебральную гемодинамику. Ионы кальция могут и прямо активировать некоторые ферменты, связываясь со специальными участками на молекулах ферментов и изменяя их конформацию, а следовательно и каталитические свойства. Среди прямых эффектов кальция большое значение имеет его влияние на активность связанного с клеточной мембранной фермента протеинкиназы С. Этот фермент, в свою очередь, фосфорилирует многочисленные белки-субстраты, стимулируя их ферментативную активность.
Читайте также:
- Статистические рассчеты фетометрии. Определение срока беременности
- Бифидобактерии и лактобактерии. Бифидумбактерин для новорожденных и детей
- Хронический средний отит с ретракционным карманом
- Диагностика врожденной мышечной дистрофии на МРТ, КТ
- Психогенный запор: случай успешного комплексного лечения