Период роста в оогенезе. Рост фолликулов
Добавил пользователь Дмитрий К. Обновлено: 22.01.2026
Кафедра акушерства и гинекологии Сибирского государственного медицинского университета Минздрава России, Томск, Россия
Сибирский государственный медицинский университет, Томск
Тканевые и молекулярные основы фолликулогенеза. Старение яичников
Журнал: Проблемы репродукции. 2017;23(4): 18‑23
Настоящий обзор научных исследований включает модели старения яичников, современные представления о фолликулогенезе и постнатальном оогенезе. Овариальная редукция непрерывно происходит в течение всей жизни женщины, однако скорость снижения овариального резерва различна в отдельные периоды жизни, что доказано с помощью математического моделирования. В результате дискоординации фолликулогенеза нарушается функционирование репродуктивной системы, что приводит к развитию гинекологических заболеваний. Физиологические функции фолликулярного развития известны, однако механизмы, контролирующие этот процесс, окончательно не установлены. Наиболее перспективными направлениями репродуктивной биологии на сегодняшний день является изучение факторов рекрутирования примордиальных фолликулов и ранних этапов фолликулярного роста, когда качество и количество женских половых клеток еще не предопределено. Основой изучения таких механизмов являются молекулярно-генетические исследования, например, создаются модели на экспериментальных животных с целенаправленными нарушениями аллелей. Следовательно, появляется возможность изучать специфические молекулы, участвующие в фолликулогенезе. Многие аспекты взаимодействия факторов фолликулогенеза требуют дальнейших исследований. Вопросы постнатального оогенеза связаны с наличием герминогенных клеток в постнатальном яичнике. Подающим надежды методом лечения бесплодия при нефункционирующих яичниках, основанном на детальном исследовании постнатального оогенеза, в будущем могут стать трансплантация аутологичных стволовых клеток или оогенез in vitro.
Самое раннее описание воспроизводства относится ко временам Гиппократа (V век до нашей эры). Считалось, что «потомство» происходит путем соединения мужского и женского эякулята (менструальная кровь). Два тысячелетия спустя Уильям Гарвей провозгласил «omne vivum ex ovo» — «все живое из яйца». В эпоху Просвещения под влиянием научной революции конца XVII века Ренье де Грааф признал, что образование яйцеклеток происходит в яичнике. На протяжении полутора веков существовало ошибочное убеждение о фолликуле как собственно женской половой клетке. В 1827 г. Карл Эрнст фон Бэр обнаружил, что ооциты млекопитающих заключены внутри фолликулов яичника. Однако первое описание фолликулогенеза у человека относится только ко второй половине XX века [1].
В настоящее время общепризнано, что яичники выполняют не только функцию воспроизводства (генерация ооцитов), но и синтезируют биологически активные молекулы, в первую очередь стероиды (в основном эстрогены и прогестины), а также факторы роста, имеющие решающее значение для регулирования гипоталамо-гипофизарно-яичниковой оси [2]. Тканевые и молекулярные основы функции яичников имеют фундаментальное значение для установления биологической роли репродуктивной системы. Понимание механизмов фолликулогенеза не только определяет патогенез различных патологических состояний, но и служит отправной точкой для применения терапевтических воздействий в клинической репродуктологии [3].
Овариальная редукция. Репродуктивная система женщины функционирует в строгом соответствии с принципом сокращения [4]. Работами E. Block (1952, 1953), T. Baker (1963) более полувека назад показано, что на 4-м месяце внутриутробного развития в яичниках плода содержится около 6—7 млн незрелых половых клеток (оогониев), окруженных слоем уплощенных гранулезных клеток, образующих примордиальный фолликулярный пул [5, 6]. В любом хронологическом возрасте подавляющее большинство (более 99%) ооцитов в яичниках присутствует в примордиальных фолликулах. Наибольшая потеря примордиальных фолликулов путем апоптоза происходит во второй половине жизни плода. К моменту рождения количество оогониев редуцируется почти в 4 раза — до 1—2 млн [7]. После рождения скорость потери фолликулов несколько замедляется. В возрасте менархе количество женских половых клеток не превышает 300—400 тыс. [8]. В течение репродуктивного периода процесс редукции не только продолжается, но и постепенно ускоряется. В возрасте 30 лет количество примордиальных фолликулов составляет только 12% (10—100 тыс.), в 40 лет — 3% (1—10 тыс.) от их значений к моменту рождения. В менопаузальный период количество фолликулов снижается менее 1000 [9]. На протяжении фертильного периода овулирует всего порядка 400 ооцитов [10].
Впервые 20 лет назад M. Faddy и R. Gosden [11] на математической модели фолликулярной динамики показали, что скорость исчезновения фолликулов увеличивается в 2 раза, когда примордиальный пул сокращается до 25 тыс. фолликулов. Разработанная модель предполагает, что женщины имеют двухфазное экспоненциальное снижение числа примордиальных фолликулов: медленное от рождения до 38 лет, а далее ускоренное. Возраст определен как критический, после которого овариальный резерв резко снижается [12]. Некоторые авторы оспорили это мнение [13, 14]. Новые данные о возможности присутствия митотически активных герминогенных стволовых клеток в яичниках половозрелых мышей не вписывались в общепринятую концепцию репродуктивной биологии млекопитающих. K. Hansen и соавт. [15] показали, что овариальное старение происходит из-за постепенного увеличения скорости атрезии в течение всего репродуктивного периода.
В настоящее время наиболее надежной иллюстрацией старения яичников признана математическая модель W. Wallace и T. Kelsey [16]. Авторами включены морфоколичественные данные о примордиальных фолликулах, демонстрирующие скорость рекрутирования, дальнейшего развития или апоптоза в яичниках женщин от зачатия до менопаузы (рис. 1).
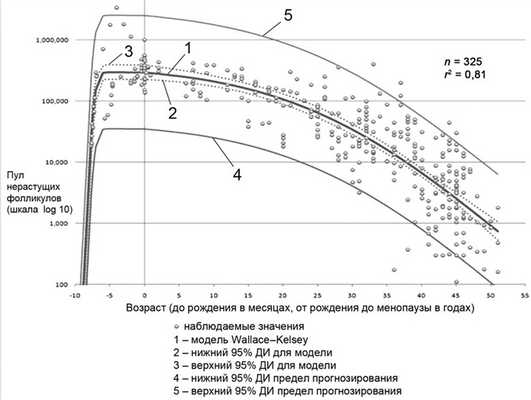
Рис. 1. Модель яичникового старения W. Wallace и T. Kelsey, 2010 [16].
Наряду с редукцией значительного количества фолликулов в течение жизни снижается качество ооцитов, по крайней мере, достигая существенной для реализации репродуктивной функции величины в возрасте старше 30 лет [5] (рис. 2). Потеря качества ооцитов обусловлена неспособностью гомологичных хромосом правильно разделиться во время мейоза, что приводит к увеличению числа анеуплоидных эмбрионов ранних этапов развития у женщин старших возрастных групп [17]. Например, риск трисомии по хромосоме 21 в возрасте 25 лет составляет 1/1500, в возрасте 40 лет — 1/16 [18]. В физиологических условиях около 20% (диапазон колебаний 10—60%) всех ооцитов формируют анеуплоидные эмбрионы. Основные механизмы могут включать повреждение герминогенных клеток при их формировании во время внутриутробной жизни, накопленные повреждения ооцитов в течение жизни женщины или возрастные изменения качества гранулезных клеток, окружающих ооцит [19].
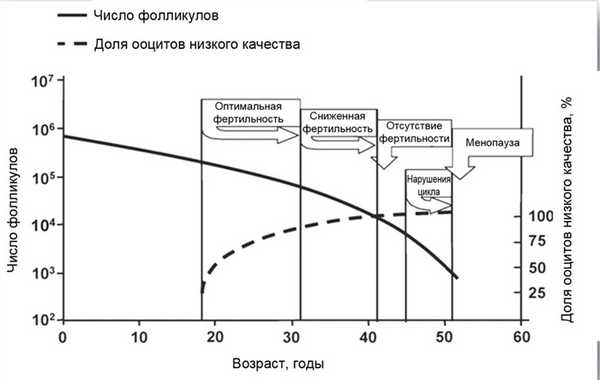
Рис. 2. Схематическое представление зависимости числа примордиальных фолликулов и компетентности ооцитов от возраста по J. de Bruin, E. teVelde (2004) [8], адаптировано F. Broekmans, M. Soules, B. Fauser (2009) [5].
На молекулярном уровне снижение качества ооцитов может быть связано с изменением биохимических процессов в митохондриях. Все митохондрии оогоний по своему происхождению материнские [18], поэтому высказано предположение, что наследование дисфункциональных старых материнских митохондрий может увеличить риск хромосомных аномалий [20]. Старение связано с увеличением митохондриальной агрегации в ооцитах и снижением концентрации церамидов [21] и цитоплазматического аденозинтрифосфата в метафазе II [22].
Еще одним «микроэкзогенным» фактором качества ооцитов является активность теломеразы. Впервые в 1971 г. отечественным ученым А.М. Оловниковым для объяснения экспериментальных данных Л. Хейфлика выдвинута теория маргинотомии — отчет клеточных делений и старения вследствие недорепликации последовательностей ДНК на концах хромосом (теломерных участков). Теория предполагала, что «нестарение» обусловлено теломеразой — особым ферментом, препятствующим укорочению теломерных участков при каждой репликации хромосом. В 1985 г. К. Грейдер и Э. Блэкберн выявили в клетках теломеразу, а в 1998 г. при помощи фермента исследователям удалось «омолодить» культуру клеток, за что в 2009 г. ученые были удостоены Нобелевской премии. Так как с возрастом активность теломеразы снижается в клетках яичников человека, несмотря на отсутствие корреляции между тандем-ДНК-полимеразой и уровнем ФСГ, активность фермента является хорошим индикатором функционального возраста яичника [23].
Фолликулогенез у млекопитающих. Фолликулогенез рассматривается как процесс достижения последовательно более высоких уровней организации с помощью пролиферации и дифференцировки клеток. После выхода из покоящегося состояния фолликулы достигают стадии овуляции либо подвергаются атрезии [24]. Развитие фолликулов включает четыре основных этапа: 1) инициацию примордиальных фолликулов; 2) ранний фолликулярный рост (развитие преантральных фолликулов); 3) отбор и рост антральных фолликулов (≥2 мм); 4) созревание преовуляторного фолликула [25]. Механизмы выхода примордиальных фолликулов из состояния покоя окончательно не установлены [26]. Преантральная стадия роста автономная (гормоннезависимая), длится наиболее долго: от инициации примордильных фолликулов до стадии роста вторичных фолликулов проходит в среднем 290 дней (~10 регулярных менструальных циклов). Антральная стадия роста возможна только в присутствии гонадотропных гормонов, прежде всего фолликулостимулирующего. После образования полости в фолликуле скорость его роста увеличивается. Интервал между кавитацией или начальными этапами образования фолликулярной полости (фолликулы диаметром 0,4 мм) до развития преовуляторного фолликула (диаметром 20 мм) составляет около 60 дней (~2 менструальных цикла). Доминантный фолликул отбирается из когорты фолликулов диаметром более 2 мм в конце лютеиновой фазы цикла. Фолликулы примерно одного диаметра (2—5 мм) формируют группы и растут под влиянием возрастающих концентраций ФСГ синхронно. Для достижения доминантным фолликулом преовуляторной стадии требуется около 20 дней. В процессе своего развития большинство фолликулов находится на пути к дегенерации и апоптозу [27—29].
Фолликулогенез происходит постоянно и безостановочно, его дискоординация может приводить к различной гинекологической патологии. Физиологические функции фолликулогенеза известны, однако механизмы, детально контролирующие этот процесс, окончательно не установлены [3]. Большинство молекулярно-генетических исследований в области репродуктивной биологии основано на изучении гомологичных генов у низших организмов и на моделях с целенаправленными нарушениями аллелей (генетически модифицированные мыши) [30]. Работы позволили не только выявить специфические молекулы, вовлеченные в фолликулогенез у человека, но также оказывались в состоянии определить мишени для потенциальных лекарственных средств в лечении патологии яичников [31]. Наиболее полная база данных генов, участвующих в «физиологическом» фолликулогенезе и при отдельных вариантах патологии гонад (синдром поликистозных яичников и преждевременное угасание функции яичников), представлена J. Hua и соавт. [3]. Интегрированная система «фолликул online» основана на более чем 43 тыс. публикаций, описывает 580 генов/белков на 23 моделях организмов, включая человека. Несмотря на большой накопленный объем информации, многие аспекты взаимодействия факторов фолликулогенеза еще предстоит изучить. Особый интерес представляют молекулярно-биологические исследования о состоянии яичников при патологических процессах, сопровождающихся вторичной овариальной недостаточностью. Длительное время основной поиск в области биологии репродукции человека был сосредоточен на последних 2 нед созревания фолликулов. Вместе с тем критически важным является понимание механизмов активации и начального фолликулярного роста, когда количество и качество ооцитов еще не определены окончательно [32].
Постнатальный оогенез. Спор о наличии или отсутствии в постнатальном яичнике герминогенных клеток возник чуть менее 100 лет назад. К середине XX века после публикации S. Zuckerman [33], показавшего, что в течение жизни большинства млекопитающих ооциты не образуются, общепризнанной концепцией считалась теория конечного количества фолликулов, которое формируется в эмбриональный период. В историческом контексте публикация J. Johnson и соавт. [13] «Герминогенные стволовые клетки и фолликулярное обновление в постнатальном яичнике млекопитающих» в журнале «Nature» подняла дискуссию на новый уровень. Авторами показано, что популяция герминогенных стволовых клеток присутствует в яичнике взрослой мыши, а число ооцитов регулируется балансом активации и гибели этих клеток.
Исследования с использованием маркировки ядер тритием тимидина показали, что ооциты постнатальных яичников являются прямыми потомками эмбриональных герминогенных клеток [33]. Незрелые половые клетки фетальных яичников млекопитающих и грызунов экспрессируют несколько факторов, характерных для плюрипотентных стволовых клеток, таких как октамерсвязывающий фактор транскрипции 4, специфический эмбриональный антиген 4, гомеобокс-фактор транскрипции генов, рецептор тирозинкиназы для фактора стволовых клеток и др. [9]. Однако не все оогонии демонстрируют аналогичную экспрессию в постнатальном периоде. Более того, после родов только у некоторых видов млекопитающих (наиболее примитивные приматы) незрелые половые клетки выявляются в гонадах. У человека почти все оогонии вступают в мейоз на 9—22-й неделе внутриутробного развития или останавливаются на стадии профазы I, элиминируются из фетальных яичников и не обнаруживаются в постнатальном периоде [34]. В целом данные свидетельствуют о примитивности неонатальных яичников мокроносых приматов по сравнению с человеческими, одновременно подтверждая факт персистенции оогониев во взрослых гонадах, учитывая эволюционное развитие приматов.
Несмотря на отсутствие прямых доказательств сохранения оогониев в яичниках взрослой женщины, идея о герминогенных стволовых клетках получила развитие [35, 36].
K. Zou и соавт. [14] успешно проведены эксперименты с герминогенными стволовыми клетками. Клетки, полученные от неонатальных мышей, трансплантировали в яичник бесплодных особей, в дальнейшем подвергались оогенезу, демонстрировали нормальный кариотип, высокую активность теломеразы и формировали потомство.
Находки в моделях на грызунах сопровождались аналогичными открытиями в организме человека. Y. White и соавт. [37] выделили митотически активные герминогенные стволовые клетки из коры яичников женщин репродуктивного возраста. Профиль экспрессии генов в них был сопоставим с примитивными половыми клетками. Авторы обнаружили, что герминогенные стволовые клетки способны сохраняться in vitro в течение нескольких месяцев и спонтанно развиваться в ооцитоподобные структуры диаметром 35—50 мкм. После маркировки зеленым флуоресцентным белком и ксенотрансплантации в биоптаты коркового вещества яичников человека или иммунодефицитных мышей наблюдалось образование «ооцитов», экспрессирующих данный белок. Следует все же отметить, что данная работа критикуется некоторыми учеными [9], учитывая потенциальные риски контаминации используемого материала незрелыми ооцитами.
Экспериментальные данные показали, что клетками-предшественниками ооцитов в постнатальных яичниках могут быть и другие популяции стволовых клеток. Работа A. Bukovsky и соавт. [38] впервые демонстрирует в качестве источника стволовых клеток поверхностный целомический эпителий [38], что находит свое подтверждение в других исследованиях [9, 39]. Ооцитоподобные клетки могут также развиваться из стволовых клеток амниотической жидкости человека, эмбриональных клеток дермы и даже панкреатических стволовых клеток в моделях на животных. Таким образом, разнообразные стволовые клетки, проявляющие плюрипотентность, могут являться предшественниками ооцитоподобных клеток, экспрессирующих зародышевые маркеры. Однако они все далеки от реальных и компетентных ооцитов.
Несмотря на ограниченные результаты исследований и большую долю скептицизма в отношении неооогенеза, направление считается весьма перспективным для репродуктивной медицины. Трансплантация аутологичных стволовых клеток и/или оогенез in vitro, вероятно, не восстановят естественную фертильность в ближайшее время, но могут вселять надежду преодоления бесплодия у пациенток с нефункционирующими яичниками. Для онкофертильности такой подход являлся бы даже более безопасным, чем криоконсервация и аутотрансплантации размороженной ткани яичников у больных раком, сопряженные с риском реимплантации злокачественных клеток в организм.
Развитие женских половых клеток — оогенез
Женская половая клетка — яйцеклетка (ovum — яйцо) — была открыта К. Бэром в 1825 г. Развитие женских половых клеток— оогенез, происходит в яичнике и завершается в яйцеводе. В оогенезе различают три стадии: размножения, роста и созревания (см. рис. 13).
Стадию размножения женских половых клеток у млекопитающих отмечают в утробный период жизни особи. Первичные половые клетки, вселившись в закладку яичника, превращаются в оогонии и много раз подряд делятся митозом — идет накопление половых клеток. Оогонии — круглые или овальные клетки с оксифильной цитоплазмой и крупным ядром, в котором
Вракин В.Ф, Сидорова М.В.
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
видны 1—2 ядрышка и мелкодисперсный хроматин. Среди соматических клеток яичника они выделяются своими крупными размерами.
Рис. 17. Рост и созревание женских половых клеток в яичнике (А)
I — корковое вещество яичника; II — мозговое вещество; III — воронка и IV — просвет яйцевода; 1 — покровный эпителий; 2 — белочная оболочка; 3— интерстициальная ткань; 4 — первичный (примордиальный) фолликул; 5
—растущий (вторичный) фолликул; 6 — пузырчатый фолликул; 7 — тека
фолликула; 8 — фолликулярный эпителий; 9 —зернистый слой; 10— полость фолликула; 11 — лучистый венец; 12 — блестящая оболочка; 13 — ооцит I порядка; 14 — овулированный фолликул; 15 — желтое тело; 16 — беловатое тело; 17 — атретический фолликул; 18 — сосуды; 19 — ооцит II порядка; 20 — зрелая яйцеклетка; 21 —полярное тельце; 22 — зигота.
К концу плодного периода у млекопитающих большинства видов митотическая активность оогониев затухает. У кроликов и свиней, по данным некоторых исследователей, размножение оогониев продолжается некоторое время после рождения (у свиней — в течение первого месяца). Прекратив делиться, половые клетки вступают в стадию роста (рис. 17).
Стадия роста оогенеза проходит в яичнике, отличается длительностью и сложностью преобразований. Половая клетка на этой стадии называется ооцитом I порядка. В его ядре происходят преобразования, характерные для профазы I мейоза. У большей части ооцитов I порядка отмечают синхронные последовательные стадии профазы I, вплоть до диплонемы. При этом половая клетка окружается одним слоем клеток — фолликулярным эпителием. Образуется многоклеточная структура — фолликул. Фолликул, стенки которого образованы одним слоем плоских фолликулярных клеток, плотно прилегающих к ооциту I порядка, называется примордиальным, а затем, когда фолликулярные клетки увеличиваются в размерах, — первичным. В первые дни после рождения ооциты проходят диплонему и вступают в про-
должительную интерфазную стадию диктиотены, предшествующую диакинезу. Она длится до полового созревания и заканчивается незадолго перед овуляцией. В это время хромосомы становятся невидимыми, их ДНК деконденсируется, активно идет синтез всех видов РНК, различных белков, увеличивается количество органелл, большинство которых располагается вокруг ядра. В результате всех этих процессов происходит рост ооцита I порядка. Рост совершается благодаря ассимиляционной деятельности половой клетки. Идет он медленно, продолжается долго— месяцы и годы и называется пе-
риодом малого, или цито-плазматического роста. Первичные фолликулы с ооцитами I порядка в стадии малого роста располагаются по периферии корковой зоны яичника. У взрослой коровы они составляют почти 55% от всех фолликулов яичника. Постепенно фолликулярные клетки увеличиваются в размерах и начинают активно функционировать. Такой фолликул называется растущим (вторичным). У коровы вторичные фолликулы составляют 43%. При совместной деятельности фолликулярных клеток и ооцита I порядка вокруг половой клетки формируется блестящая оболочка — вторичная оболочка половой клетки. Через ее поры устанавливается тесная связь половой и фолликулярных клеток. Последние, размножаясь, покрывают ооцит I порядка несколькими слоями. Особенно активен фолликулярный эпителий в период накопления желтка — в период вителлогенеза (vitellus — желток, genesis — происхождение), который начинается при половом созревании. С помощью фолликулярного эпителия идет быстрое накопление ооцитом I порядка бел-
лина), синтезированных за пределами яичника, в основном в печени и принесенных кровью.
Период вителлогенеза называют еще периодом большого роста, или трофоплазматическим. У свиньи он протекает в фолликулах с диаметром от 4 до 8 мм. В это время происходит пропорциональное увеличение ооцита I порядка и фолликула. Фолликулярные клетки осуществляют отбор и концентрацию вокруг ооцита молекул, подлежащих эндоцитозу. Они же, повидимому, выделяют вещества, стимулирующие микропиноцитоз в ооците и гормоны, стимулирующие синтез вителлогенина печенью. Живые неокрашенные ооциты I порядка в период большого роста имеют коричневый цвет. Органеллы в них располагаются в основном по периферии цитоплазмы, где в первую очередь идет отложение желточных гранул и других включений. Постепенно желток заполняет большую часть цитоплазмы ооцита, вителлогенез затухает. Рост ооцита прекращается, а фолликул продолжает расти.
Между фолликулярными клетками появляется полость, в которой накапливается жидкость, содержащая половые гормоны (эстрогены) и другие продукты деятельности фолликулярного эпителия. Фолликул становится пу-
зырчатым (Граафов пузырек или третичный фолликул). У коровы полость начинает появляться в фолликулах, достигших диаметра 1 мм. Место, где в стенке фолликула расположен ооцит I порядка, называется яйценосным бугорком. Ооцит покрыт слоем фолликулярных клеток или лучистым венцом, который у сельскохозяйственных животных образован 8—12 рядами клеток.
Остальные фолликулярные клетки формируют зернистый слой фолликула— гранулезу. Поверх него фолликул одет соединительнотканной оболочкой — текой. Тека выполняет опорную и трофическую функции, в ней много сосудов и нервов, залегают клетки, вырабатывающие половые гормоны. Созревший пузырчатый фолликул имеет макроскопические размеры (табл. 2).
2. Размеры женских половых клеток и фолликулов на разных стадиях развития (по данным разных авторов)
Яйцеклетка с оболочками
Ооцит в профазе мейоза
Незадолго до конца стадии роста хромосомы в ядре ооцита I порядка конденсируются и принимают вид, характерный для диакинеза профазы I, после чего мейоз идет без остановки.
В определенный момент полового цикла под влиянием нейроэндокринной регуляции расширяются кровеносные сосуды теки, резко возрастает давление жидкости внутри фолликула. Стенка его разрывается, и половая клетка вместе с лучистым венцом выпадает в яйцевод. Этот процесс называется овуляцией.
Стадия созревания у большинства млекопитающих начинается в яичнике и завершается (с метафазы II) в яйцеводе. Исключение составляют лошадь, собака, у которых половая клетка овулирует стадии диакинеза профазы I. В стадии созревания половая клетка делится мейозом. В результате первого деления созревания— редукционного деления мейоза из одного ооцита I порядка— образуются две неравнозначные клетки. Одна — крупная половая клетка, содержит всю цитоплазму и половину хромосом. Это ооцит II порядка. Другая — мелкая, содержит половину хромосом и почти не имеет цито-
плазмы, первое редукционное, или полярное, тельце. Сразу после завершения первого деления созревания наступает второе деление созревания — эквационное деление мейоза, результатом которого вновь являются две неравно-
ценные клетки: зрелая яйцеклетка и второе редукционное тельце. Иногда
первое редукционное тельце также делится эквационно. Импульсом для деления созревания у многих животных является совокупление или проникновение спермия в оболочки яйцеклетки. Если этого не произойдет, половая клетка гибнет на стадии метафазы II. Все редукционные тельца в дальнейшем отмирают. Таким образом, в оогенезе из одного ооцита I порядка образуется лишь одна, но очень крупная половая клетка — яйцеклетка, содержащая гаплоидный набор хромосом и полный набор факторов, необходимых для начальных этапов развития зародыша.
Строение зрелой яйцеклетки. Яйцеклетка, как и любая клетка, состоит из ядра, цитоплазмы и цитолеммы. В то же время по своим свойствам и структуре она коренным образом отличается от всех других клеток организма.
Ядро яйцеклетки небольших размеров, неактивное, с гаплоидным набором хромосом. Ядерно-цитоплазматическое отношение сильно сдвинуто в сторону преобладания цитоплазмы. В цитоплазме яйцеклетки происходит развитие морфогенетической организации, формирование специализированной кортикальной зоны. В ней накоплены разнообразные вещества, необходимые для ранних стадий развития зародыша: все виды РНК, многообразные ферменты, структурные белки, энергетические резервы, множество факторов, выполняющих регуляторные функции, благодаря деятельности которых как бы развертывается план строения раннего зародыша. Особенно в больших количествах накапливается желток, в состав которого входят белки (вител-
вителлин и др.). 80% белков цитоплазмы яйцеклетки приходится на долю желтка, 16% составляют структурные белки рибосом.
Размеры яйцеклетки определяются количеством желтка. Количество желтка зависит от условий и длительности развития зародыша. У животных с личиночным (беспозвоночные), коротким (ланцетник) и внутриутробным (млекопитающие) развитием яйцеклетки содержат мало желтка и называются олиголецитальные. У животных с прямым наружным развитием (рептилии, птицы) яйцеклетки полилецитальные, в них много желтка. Если желток расположен в яйцеклетке равномерно, она относится к изолецитальному типу (ланцетник), и если скапливается у одного полюса — к телолецитальному типу (птицы). Олиголецитальные яйцеклетки гораздо меньше полилецитальных. Масса яйцеклеток у млекопитающих в среднем 3×10 -6 г, диаметр 60— 180 мкм, у ланцетника — 100 мкм, у домашних птиц — 30—50 мм, у страуса— 105 мм, а у одного вида акул — до 220 мм.
Вещества и органеллы цитоплазмы яйцеклетки не разбросаны в беспорядке, а организованы столь определенным образом, что эта структурная организация во многом определяет последующие этапы эмбрионального развития. Вот один пример. Полюс, у которого скапливается желток, называется вегетативным. Ядро яйцеклетки при этом располагается у противоположного полюса— анимального. Данными экспериментальной эмбриологии установлено, что оси будущего зародыша определяются этой полярностью. Анимальный полюс становится головным концом зародыша, вегетативный —
хвостовым. По особенностям структурной организации разных участков цитоплазмы яйцеклетки можно определить брюшную и спинную стороны будущего зародыша, плоскость билатеральной симметрии и другие показатели.
В периферическом слое цитоплазмы ооцита, или в кортикальном, располагается большое количество микрофибрилл, делающих его вязким. Здесь же скапливаются так называемые кортикальные гранулы, или кортикальные вакуоли, которые содержат гликозаминогликаны. Кортикальный слой цитоплазмы, формирующийся еще в период роста ооцита, играет важную роль при оплодотворении.
Яйцеклетка покрыта оболочками. Первичная оболочка синтезируется самим ооцитом в период роста и называется желточной, или вителлиновой. Она тонким слоем покрывает ооцит, располагаясь между его микроворсинками. К ней тесно прилегает вторичная оболочка, образованная деятельностью фолликулярных клеток. У млекопитающих первичная и вторичная оболочки столь тесно прилежат и пронизывают друг друга, что известны под общим названием блестящей, или прозрачной, оболочки. Строение ее у сельскохозяйственных животных сходно, толщина 10—: 20 мкм. Она является мощным иммунным барьером, обладает избирательной проницаемостью для спермиев. Третичная оболочка продуцируется железами яйцевода и покрывает яйцеклетку, когда она движется по яйцеводу. Третичная оболочка может иметь сложное строение. У птиц, например, она состоит из белка, нодскорлуповой, скорлуповой и надскорлуповой оболочек, образующихся в разных
Влияние различных факторов на оогенез. В процессе оогенеза на всех стадиях развития происходит гибель какой-то части половых клеток под влиянием внутренних, во многом не выясненных, факторов. Отмечено три волны массовой дегенерации: на стадии размножения оогонии, ооцитов I порядка в стадии пахитены и в стадии диплотены. В результате число половых клеток к рождению уменьшается в 7—10 раз. Гибель клеток (так называемая атрезия) продолжается и в дальнейшем. Считается нормальным, если из 20 растущих фолликулов до овуляции дорастает один. У коров критическим моментом в стадии роста является также период достижения фолликулом размеров от 6 до 9 мм. В это время происходит атрезия 55% пузырчатых фолликулов. По-видимому, так же как в сперматогенезе, основная причина гибели половых клеток в оогенезе — нарушения в хромосомах и веретене деления. В результате оогония или ооцит теряют способность к дальнейшему развитию и погибают. К атрезии фолликулов ведет и нарушение гормональных взаимоотношений (между эстрадиолом, тестостероном и прогестероном) в процессе оогенеза.
Большинство повреждающих внешних факторов (высокая или низкая температура, действие химических веществ, электрошок) нарушает течение митоза или мейоза в результате разрушения веретена деления. Ионизирующая радиация и некоторые химические вещества воздействуют непосредственно на хромосомы. Погибают половые клетки или образуются их атипичные формы. Следствием этого может быть бесплодие, снижение пло-
Период роста оогенеза: процессы, происходящие в фолликуле и ооците 1 порядка.
Стадия роста оогенеза протекает в яичнике. Она делится на два периода. Период малого, или цитоплазматического роста протекает от рождения до начала полового созревания. Второй период большого, или трофоплазматического роста, начинается при половом созревании и продолжается до конца репродуктивного периода женской особи. Половая клетка в стадию роста называется ооцит I порядка. Сначала в ядре молодой яйцеклетки происходят изменения характерные для профазы I мейоза. Ооциты I порядка в яичниках образуют структуры называемые фолликулами. Различают четыре типа фолликулов. Примордиальный фолликул состоит из ооцита I порядка, окружаенного одним слоем плоских фолликулярных клеток. Первичный фолликул образован ооцитом I порядка, окруженным слоем высоких фолликулярных клеток. Вторичный фолликул представлен растущим ооцитом I порядка, в цитоплазме которого идет активное накопление питательных веществ. Он окружен блестящей оболочкой и несколькими слоями фолликулярных клеток. Третичный фолликул, или граафов пузырек - окружен соединительнотканной оболочкой - текой и зернистым слоем гранулезой, содержит жидкость, имеет на стенке яйценосный бугорок, на котором располагается молодая яйцеклетка на стадии ооцита I порядка. Вокруг нее фолликулярные клетки формируют лучистый венец. В определенный момент полового цикла под влиянием гормонов стенка пузырчатого фолликула лопается и половая клетка с лучистым венцом попадает в воронку яйцевода. Процесс выхода яйцеклетки из яичника называется овуляция.
Общие и отличительные черты сперматогенеза и оогенеза.
Гаметогенез - это процесс образования и развития гамет. Различают сперматогенез - процессы образования сперматозоидов и оогенез (овогенез) - процесс образования яйцеклеток. У животных гаметогенез происходит в половых железах - семенниках и яичниках, а у растений в специальных органах - гаметангиях. Гаметы развиваются из клеток-предшественников (гоноцитов), которые диплоидны.
Различают 4 периода гаметогенеза:
Период размножения. Клетки-предшественники делятся путём митоза, количество клеток увеличивается. Клетки остаются диплоидными
Период роста. Происходит рост клеток, количество цитоплазмы увеличивается. Клетки находится в состоянии интерфазы.
Период созревания Во время периода созревания происходит 2 деления мейоза, в результате количество хромосом уменьшается в 2 раза, набор хромосом становится гаплоидным.
Период формирования (бывает только в сперматогенезе). Сперматозоид приобретает специфические особенности строения
Этапы оплодотворения и его биологическое значение.
ОПЛОДОТВОРЕНИЕ - сложный комплекс физиологических процессов и реакций, приводящий к слиянию мужской и женской половых клеток в одну клетку - зиготу (zygotes - объединенный). Оно протекает в четыре стадии.
сближение половых клеток;
проникновение спермиев в блестящую оболочку;
проникновение в цитоплазму яйцеклетки;
слияние ядер половых клеток.
В половые пути самок попадают миллиарды спермиев, которые движутся благодаря реотаксису (способности двигаться против тока жидкости) и присасывающего действия мускулатуры матки и яйцеводов. Через 7-9 часов спермии оказываются в верхней трети яйцевода. Доходит туда только 1% спермиев. Основная их масса гибнет в пути. При контакте яйцеклетки со спермиями происходит акросомная реакция. Из акросомы спермия освобождаются ферменты, которые разрушают прозрачную оболочку и лучистый венец яйцеклетки. Спермии проходят через блестящую оболочку, плазматические мембраны спермия и яйцеклетки вступают в соприкосновение и сливаются. Это служит толчком для пробуждения яйца. С этого момента снимается блокада с мейоза и он продолжается с той стадии, на которой остановился во время овуляции. Завершается первое и начинается второе деление мейоза. Резко активизируется обмен веществ в яйцеклетке. Развивается кортикальная реакция. Кортикальные гранулы цитоплазмы подходят к плазмолемме, изливаются в сторону прозрачной оболочки. При этом образуется превителлиновое пространство, в результате чего яйцеклетка отслаивается от прозрачной оболочки, которая стабилизируется и преобразуется в оболочку оплодотворения. Она препятствует проникновению других спермиев. У олиголецитальных яйцеклеток только один спермий попадает внутрь, это называется моноспермия. У полилецитальных животных наблюдается полиспермия - несколько спермиев проникают внутрь яйцеклетки. И в том, и в другом случае с ядром яйцеклетки сливается только один спермий. Ядра половых клеток (пронуклеусы) набухают, мигрируют к центру яйцеклетки и сближаются. Их ядерные оболочки исчезают, материнские и отцовские хромомсомы объединяются. Образуется одноклеточный зародыш зигота.
Ооциты и оогенез
Наш организм состоит из множества различных клеток, которые формируют органы и ткани. Все они имеют свои индивидуальные особенности, но в целом похожи друг на друга. Самые большие и долгоживущие клетки в организме, имеющие существенные отличия от других - это ооциты. Они являются незрелыми яйцеклетками, которые начинают продуцироваться уже во внутриутробном периоде.
Ооциты: что это такое
Ооциты - это основные элементы репродуктивной женской системы, которые дают возможность зарождения новой жизни. Можно сказать, что они являются одной из стадий развития яйцеклеток. Выделяют ооциты первого и второго порядка.
Ооциты первого порядка - это клетки, которые образовались в результате митотического деления первичных половых клеток (оогоний) и накопили достаточное количество питательных веществ, они появляются во внутриутробном периоде и остаются неизменными вплоть до полового созревания (12-15 лет). К моменту рождения в яичниках содержится порядка 2 миллионов ооцитов, большинство из которых никогда не продолжат дальнейшее развитие до полноценной яйцеклетки и погибнут. Все процессы деления пройдут лишь 400-500 ооцитов. Для чего необходима столь избыточная продукция половых клеток и почему значительное их количество погибает, до сих пор неизвестно.
Ооциты второго порядка - это клетки, которые образовались в результате завершения 1 профазы мейоза ооцитов первого порядка. Мейоз - это процесс деления половых клеток, который делает их гаплоидными (они содержат только 1 набор хромосом). 1 профаза мейоза начинается после полового созревания и стимулируется различными факторами, в частности половыми гормонами.
В 1 профазу мейоза одновременно вступает порядка 15000-20000 ооцитов первого порядка, но завершает процесс только 1 клетка. Все остальные редуцируются и погибают. Затем ооциты второго порядка вступают во 2 профазу мейоза и формируют зрелую яйцеклетку, готовую к оплодотворению.
Ооциты второго порядка, которые формируются в конце репродуктивного периода, могут быть неполноценными и дефектными. Ведь они образуются из ооцитов первого порядка, живущих примерно 40-50 лет, а в процессе их существования могут происходить разнообразные генетические сбои и другие неблагоприятные явления. Поэтому число случаев аномалий развития у детей, рожденных после 40 лет, значительно увеличивается.
Оогенез
Оогенез - процесс развития женских половых клеток. Он принципиально отличается от сперматогенеза - процесса формирования мужских половых клеток. Конечно, сперматозоиды и ооциты (яйцеклетки) образуются в результате митоза и мейоза первичных зародышевых клеток, но при этом сперматозоиды являются высокодифференцированными клетками, которые в основном содержат лишь генетический материал, а яйцеклетки, помимо генов, включают все необходимые элементы для эмбрионального роста. Таким образом, ооциты - это большие малодифференцированные клетки, состоящие из множества структурных белков, ферментов и питательных веществ.
Помимо этого, есть количественные различия между процессами гаметогенеза, ведь мужчина продуцирует порядка 30 миллионов сперматозоидов в сутки, а ооциты закладываются еще до рождения, при этом зрелыми яйцеклетками становятся только ~500 из них.
Первичные половые клетки - оогонии - попав в яичники, начинают активно размножаться путем митоза и формируют примордиальные фолликулы и ооциты первого порядка. Затем они подвергаются процессу мейоза: в первой фазе образуются ооциты второго порядка, в конце 2 - зрелые и полноценные яйцеклетки, которые выходят из фолликула и происходит овуляция.
Зрелая клетка содержит большой запас питательных веществ и микроэлементов, необходимых для развития эмбриона в первое время. Размеры яйцеклеток отличатся от других клеток организма, у людей они достигают 60-2000 мкм, у рыб - 6-9 мм, у страуса - несколько сантиметров.
Этапы оогенеза
Оогенез делится на несколько последовательных этапов.
Первый этап - это перемещение первичных половых клеток в гонады и их размножение. Первоначально образуется несколько десятков половых клеток, но по мере их продвижения в яичники они активно делятся и их число возрастает до нескольких тысяч. В гонадах они также продолжают размножаться путем митоза, в результате к 7 месяцу внутриутробного развития насчитывается порядка 7 млн. оогоний. Но большая часть погибает еще до рождения, поэтому их остается лишь 2 млн.
Клетки, которые завершили деление путем митоза, вступают в новый этап оогенеза. Теперь они называются - ооциты первого порядка. С момента рождения девочки они остаются без изменений вплоть до полового созревания. Стоит отметить, что большая часть ооцитов никогда не вступают в дальнейший этап и только 400-500 завершают свое развитие. Еще во время эмбрионального развития некоторые ооциты первого порядка вступают в первую профазу мейоза, но не завершают ее до полового созревания. Они накапливают питательные вещества и увеличиваются в размерах. При этом они покрываются слоем фолликулярных клеток, образуя примордиальные фолликулы.
Второй этап - первая профаза мейоза, в результате которой формируется ооцит второго порядка и редукционное тельце. Под действием половых гормонов завершается процесс, который начался еще до рождения. В результате деления происходит неравномерное разделение цитоплазмы, большая ее часть остается в ооците, а редукционное тельце содержит в основном только ядро. Ооциты второго порядка обладают гаплоидным набором хромосом.
Третий этап - вторая профаза мейоза, которая приводит к формированию полноценной яйцеклетки, готовой к оплодотворению. Ооциты второго порядка вновь неравномерно делятся, образуется яйцеклетка и редукционное тельце, первое тельце также разделяется. В итоге после завершения процесса оогенеза формируется 3 редукционных тельца и яйцеклетка.
Качество ооцитов и как его улучшить
Ооциты должны содержать большое количество питательных веществ и гаплоидный набор хромосом для осуществления зачатия. Часто женское бесплодие связано с низким качеством ооцитов. На сегодняшний момент времени не разработано эффективного лечения по устранению этого фактора. Хотя женщинам в период планирования беременности рекомендуется принимать мультивитамины, в частности фолиевую кислоту и витамин Е, морепродукты, рыбий жир, которые необходимы для полноценного эмбрионального развития. Также следует отказаться от алкоголя и курения, кофе, жареного, черного чая, наладить ритм жизни, соблюдать режим труда и отдыха.
Многие специалисты считают, что окислители и свободные радикалы могут существенно снизить качество ооцитов. С целью предотвращения негативного воздействия назначают мелатонин, который противодействует окислительному процессу.
Ооциты и их доноры
В наше время для многих последняя надежда на долгожданную беременность - вспомогательные репродуктивные технологии: ЭКО, ИКСИ и другие. Но иногда возникают ситуации, когда у женщины невозможно получить полноценные яйцеклетки:
- Менопауза или раннее ее наступление;
- Неразвитые гонады и их отсутствие;
- Химиорадиотерапия;
- Наследственные заболевания, передающиеся по женской линии;
- Множественные неудачные попытки ЭКО.
Единственной возможностью для женщины самостоятельно выносить ребенка - обратиться к донору ооцитов. К донорам предъявляются довольно жесткие требования: они должны быть соматически, физически и психически здоровы. В некоторых европейских странах такой метод лечения запрещен в силу этических и религиозных соображений.
Иногда женщина, согласившись передать свои ооциты, не знает - теряет ли она что-то безвозвратно. Но в случае донорства используются клетки, которые никогда не будут использованы во время жизни, следовательно, процедура не приводит к уменьшению собственных запасов. Часто бесплодные пары приводят своих доноров ооцитов: ими могут быть близкие родственники, друзья или знакомые. Но у всех есть возможность обратиться в банк доноров, в них включаются женщины в возрасте от 18 до 35 лет, у которых уже есть ребенок, они полностью дееспособны и прошли полное медицинское обследование. Также в этом банке можно подобрать донора с определенными внешними признаками: цвет глаз, рост, телосложение и другие.
Следует отметить, что многие женщины, воспользовавшись донорскими ооцитами, после приживления эмбрионов и наступления беременности забывают, что генетически ребенок имеет родство только с мужчиной. Но к этому решению следует приходить обдуманно, взвесив все достоинства, недостатки и обсудив с супругом.
Пройти полное обследование, сдать необходимые анализы и получить ответы на интересующие вас вопросы вы можете в клинике «Центр ЭКО» в Смоленске.
Процесс формирования и строение зрелого спермия.
Стадия формирования. Сперматиды превращаются в спермии или сперматозоиды. Все сперматиды одного клона одновременно проходят эту стадию. В процессе формирования на переднем полюсе сперматиды из комплекса Гольджи образуется акросома - специфическая органелла, содержащая ферменты (преимущественно гиалуронидазу) и обеспечивающая растворение оболочек яйцеклетки. Она примыкает к ядру и по мере развития прикрывает его. Ядро вытягивается, уплотняется в 20-50 раз, его хромосомы конденсируются. От центриоли отрастает жгутик, направленный к просвету канальца. Часть цитоплазмы фагоцитируется клеткой Сертоли. В конце стадии формирования из сперматиды образуется спермий.
Строение спермия. Спермий - свободная подвижная клетка, длиной у млекопитающих 40-60 мкм. Состоит из головки, шейки и хвоста. Все его части покрыты цитолеммой. Головка имеет овальную уплощенную форму, на 2/3 покрыта акросомой. Основу головки составляет ядро, в котором содержится ДНК в виде плотно упакованных хромосом. Цитоплазма очень тонким слоем покрывает ядро. Шейка - самый короткий отдел, видимый только в электронный микроскоп. В ней располагается центросома из двух центриолей. Проксимальная центриоль лежит ближе к ядру поперек шейки, в дистальная, наиболее удаленная, лежит вдоль шейки. От нее берет начало осевая нить (аксонема) жгутика. Хвост составляет основную длину клетки. Он состоит из среднего (промежуточного), главного и концевого отделов. Осевая нить проходит вдоль всего хвоста, состоит из микротрубочек, построенных из белков - тубулина и динеина, способных к сокращению. Благодаря им хвост спермия осуществляет волнообразные и бичевидные движения. В среднем отделе хвоста расположены митохондрии, упакованные в виде спирали (спиральная нить спермия). Средний отдел от главного отделяется замыкающим кольцом. Цитоплазма тонким слоем покрывает весь хвост, за исключением концевого отдела, иногда образует 1-2 выроста - ундулирующую мембрану, дополнительное приспособление для передвижения.
Оогенез.
ООГЕНЕЗ - развитие женских половых клеток. Женская половая клетка - яйцеклетка (ovarium - яйцо) - была открыта К. Бэром в 1825 году. Развитие женских половых клеток называется оогенез, он происходит в яичнике и завершается в яйцеводе. В оогенезе различают три стадии: размножения, роста, созревания.
Стадия размножения. Происходит во внутриутробный период жизни особи. Первичные половые клетки заселяют закладку яичников и превращаются в оогонии. Оогонии многократно делятся митозом и заполняют весь яичник. Это круглые или овальные клетки с крупным ядром, в котором видны 1-2 ядрышка и мелкодисперсный хроматин. К концу плодного периода митотическая активность оогониев затухает. У большинства домашних животных к моменту рождения стадия размножения заканчивается и наступает следующая стадия - роста.
Стадия созревания. Начинается в яичнике и заканчивается в верхней трети яйцевода. Половая клетка делится мейозом. После первого - редукционного деления из одного ооцита I порядка образуется две неравнозначные клетки. Одна - ооцит II порядка крупная половая клетка. Она содержит всю цитоплазму и гаплоидное число хромосом. Другая - мелкая клетка почти не имеет цитоплазмы и содержит половину хромосом, называется полярное, или первое редукционного тельце. После второго эквационного деления мейоза появляется зрелая яйцеклетка и второе редукционное тельце. Импульсом для деления созревания у многих животных является совокупление или проникновение в оболочки яйцеклетки. Если этого не произойдет, яйцеклетка гибнет на стадии метафазы II. Таким образом, в оогенезе из одного ооцита I порядка образуется только одна яйцеклетка с истинно гаплоидным набором хромосом.
Строение яйцеклетки. Яйцеклетка состоит из ядра, цитоплазмы и плазмолеммы. Ядро клетки небольшого размера. Цитоплазма содержит желток, в состав которого входят белки (вителлин), углеводы и липиды. На долю желтка приходится 80% белков цитоплазмы и 16% составляют структурные белки. Полюс, у которого скапливается желток, называется вегетативным. Ядро находится на противоположном - анимальном полюсе, который станет головным концом зародыша. Цитоплазма яйцеклетки формирует кортикальную зону, где накапливаются многообразные белки, ферменты, энергетические запасы, микрофибриллы и образуются кортикальные гранулы. Кортикальный слой играет важную роль при оплодотворении.
Яйцеклетка покрыта плазмолеммой, прозрачной оболочкой и лучистым венцом. Прозрачная, или блестящая оболочка образуется по мере развития яйцеклетки, содержит густую сеть тонких нитей, состоящих из гликопротеинов. Один из них формирует рецептор сперматозоида. Связывание сперматозоида с этим рецептором является сигналом к акросомной реакции. У птиц по мере прохождения яйцеклетки по яйцеводу формируются третичные оболочки, к которым относятся белочная, подскорлуповая, скорлуповая и надскорлуповая. Они образуются в разных отделах яйцевода.
Типы яйцеклеток. В зависимости от количества желтка и от его распределения в цитоплазме яйцеклетки бывают разных типов. У животных с личиночным (беспозвоночные), коротким (ланцетник) и внутриутробным (млекопитающие) развитием яйцеклетки содержат мало желтка и называются олиголецитальные. У животных с наружным прямым развитием (рептилии и птицы) яйцеклетки полилецитальные с о значительными запасами желтка. Если желток расположен в цитоплазме равномерно, она относится к изолецитальному типу (ланцетник), а если скапливается у одного полюса - к телолецитальному типу (птицы). Олиголецитальные яйцеклетки меньше по размеру, чем телолецитальные. Диаметр у млекопитающих 60-180 мкм, а у птиц - 30-50 мм.
Влияние различных факторов на оогенез. В процессе оогенеза на всех стадиях развития происходит гибель части половых клеток. В результате число половых клеток к рождению уменьшается в 7-10 раз. Гибель клеток - атрезия продолжается и в дальнейшем. Основная причина - это нарушения в хромосомах, в веретене деления, нарушения гормональных взаимоотношений между эстрадиолом, тестостероном и прогестероном в процессе оогенеза. Большинство повреждающих внешних факторов (высокая или низкая температура, действие химических веществ) также нарушают течение митоза или мейоза. Следствием этого может быть бесплодие, снижение плодовитости, повышенная эмбриональная смертность и появление уродств у плодов.
Читайте также: