Петля Генле. Нисходящий ( тонкий ) отдел петли Генле. Дистальный отдел канальцев ( дистальные канальцы ). Собирательные трубки.
Добавил пользователь Дмитрий К. Обновлено: 28.01.2026
В процессе осмотического концентрирования мочи принимают участие: петля Генле, дистальный каналец, собирательная трубка, сосуды и интерстиций мозгового вещества, которые функционируют как единая поворотно - противоточно-множительная система. Мы уже упоминали, что в концентрировании мочи участвуют юкстамедуллярные нефроны с длинными петлями Генле, которые, как и собирательные трубки, глубоко проникают в мозговой слой почки.
Сразу «раскроем карты» - процесс окончательного концентрирования мочи происходит в собирательных трубках, а условия для этого создаются работой всей поворотно - противоточно-множительной системы. Эта система создает гиперосмолярность мозгового вещества и при действии антидиуретического гормона (АДГ) заставляет воду переходить из собирательной трубки в интерстиций, а затем в кровеносные сосуды мозгового вещества, в результате чего образуется концентрированная моча. Ключевым вопросом при осуществлении концентрирования мочи является вопрос о том, каким образом интерстициальная жидкость мозгового вещества становится гиперосмолярной. Попытаемся ответить на него. Следите за нашими рассуждениями по рис.10: из проксимального канальца жидкость, изоосмотическая плазме крови, с концентрацией 300 мосм/л ,поступает в тонкое нисходящее колено петли Генле и, продвигаясь по нему, начинает терять воду; в результате ее осмотическая концентрация прогрессивно нарастает и на изгибе петли в сосочке достигает своего максимума (1400мосм/л). Затем она поворачивает и течет по восходящему колену в противоположном направлении (отсюда название поворотно-противоточная система), при этом происходит ее разбавление и уменьшение осмолярности до 100 мосм/л.Эти процессы обусловлены разными функциональными свойствами стенок этих участков петли: «маховиком» этой системы является толстый восходящий отдел петли, который совершенно непроницаем для воды, но активно реабсорбирует Nа и Cl. В апикальной мембране клеток этого отдела транспорт Nа осуществляется одновременно с ионами К и двумя ионами Cl с помощью Nа, К,2Cl, котранспортера. Через базальную мембрану клеток Nа переносится активно.
NаCl поступает в интерстиций и обуславливает выход воды из нисходящего колена петли, стенки которого, напротив, обладают высокой проницаемостью для воды, но не пропускает ионы Nа. Таким образом, реабсорбция хлорида натрия восходящей частью петли “отвечает” за реабсорбцию воды в нисходящей части. Давайте разберем это сначала на простой схеме (рис.11).
Представим себе, что петля Генле заполнена неподвижной жидкостью, которая поступила из проксимального канальца, тогда осмотическая концентрация в любой части петли Генле равна 300 мосм/л. (1). Теперь допустим (2), что активная транспортная система в восходящей части петли осуществляет реабсорбцию хлорида натрия в интерстициальное пространство до тех пор, пока не установится предельный градиент (скажем 200 мосм/л) между жидкостью, находящейся в восходящем колене петли Генле и жидкостью интерстициального пространства. Вспомните, что стенки этой части петли непроницаема для воды.
Обратите внимание на то, что теперь существует разница в осмолярности между жидкостью в нисходящем колене (300 мосм/л) и окружающей каналец интерстициальной жидкостью (400 мосм/л). Поскольку стенки нисходящего колена петли хорошо проницаемы для воды, вода пассивно выходит из его просвета в интерстициальное пространство, в котором существует более высокая осмолярность, возникшая вследствие реабсорбции хлорида натрия в восходящем колене, и осмолярность жидкости в нисходящем колене увеличивается до 400 мосм/л. (3).
Теперь усложним схему: пусть жидкость в петле не стоит на месте, а непрерывно движется, тогда по мере продвижения вниз по нисходящему колену из нее уходит все больше воды, концентрация внутриканальцевой жидкости все больше увеличивается и, поскольку устанавливается осмотическое равновесие, концентрация интерстициальной жидкости увеличивается на ту же величину (200 мосм/л). Градиент в 200 мосм/л, так называемый поперечный или горизонтальный градиент, поддерживается на каждом из «этажей» мозгового вещества.
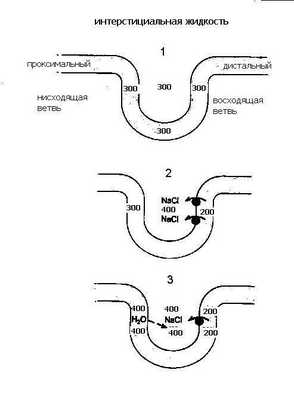
Рисунок 11 Схема взаимодействия нисходящего и восходящего колена в процессе концентрирования канальцевой жидкости.
Таким образом, между соседними участками нисходящего и восходящего колена разность осмотической концентрации невелика, но по ходу петли и длине почечного сосочка - по вертикали - этот одиночный эффект нарастает, суммируется (умножается), и в результате формируется значительно больший перепад осмотического давления - так называемый корково-сосочковый вертикальный осмотический градиент.
1400 мосм/л сосочек
Теперь Вам понятно, почему система называется ещё и множительная. Таким образом, нисходящее и восходящее колено петли тесно соприкасаются друг с другом, расположены параллельно, оказывают друг на друга влияние и функционируют как единый сопряженный механизм - противоточно-поворотно-множительная система. Благодаря работе этой системы в интерстиции мозгового вещества создается корково-сосочковый осмотический градиент.
Следует отметить, что осмолярность интерстиция создается не только ионами хлорида натрия. Примерно половина осмолярности обусловлена присутствием в нем мочевины. У мочевины имеется свой кругооборот в почке, в частности из собирательной трубки она пассивно диффундирует в интерстиций мозгового вещества и тем самым, увеличивает его осмолярность.
Прямые сосуды мозгового вещества, подобно коленам петли Генле, тоже образуют поворотно-противоточную систему. Они расположены параллельно петлям Генле и в них происходят такие же изменения осмолярности, что и в петле. Это сохраняет продольный осмотический градиент в мозговом веществе, не позволяя ему вымываться.
Вернемся к нашим рассуждениям.
Приступая к изучению работы концентрирующего механизма почки, мы поставили перед собой задачу выяснить, каким образом в интерстиции мозгового вещества создается зона гиперосмолярности, и решили её. Но пока мы не решили проблему концентрирования мочи, которую поставили перед почкой: по мере того, как канальцевая жидкость двигалась по нисходящему колену, ее осмолярность нарастала и в районе изгиба достигала 1400 мосм/л, так же как и осмолярность интерстиция, но по мере движения по восходящему колену она вновь разбавилась до 100 мосм/л. Далее в дистальном канальце происходит реабсорбция NаCl, воды и др. веществ и осмотическая концентрация вновь становится - 300 мосм/л, но по-прежнему канальцевая жидкость изосмотична крови,т.е.,в петле концентрирование не произошло. А где же будет образовываться концентрированная моча?
Помните, мы вначале “раскрыли карты”. Да, решающим обстоятельством станет то, что из дистального канальца жидкость поступает в собирательную трубку, где и будет происходить формирование окончательной мочи и процесс ее концентрирования. Неслучайно собирательные трубки расположены параллельно петлям Генле и прямым сосудам. Они транзитом проходят через все зоны почки и на всем протяжении окружены интерстицием с прогрессивно нарастающим в направлении от коры к сосочку осмотическим давлением. Обратите внимание на рисунок 10, осмолярность интерстициальной жидкости на каждом уровне идентична этой величине в нисходящем колене и СТ. Другими словами вокруг СТ на каждом “этаже” мозгового вещества имеется горизонтальный осмотический градиент в 200 мосм/л, а по вертикали - мощный корково-сосочковый осмотический градиент, созданный поворотно - противочной системой петли Генле. Таким образом, можно сказать, что петля Генле «работает» на собирательную трубку, создавая в интерстиции мозгового вещества зону гиперосмии. Это и будет та сила, которая способна вытянуть воду из собирательной трубки и произвести концентрирование мочи. Когда канальцевая жидкость поступает в собирательную трубку, ее осмолярность находится на том же уровне, что и осмолярность интерстициальной жидкости в этой зоне почки. В районе сосочка величина осмолярности достигает максимума (у человека она равна 1400 мосм/л), поэтому максимальная осмотическая концентрация мочи у человека тоже может достигать 1400 мосм/л.
Резюмируя вышеизложенное, можно представить следующую схему основных процессов, обеспечивающих осмотическое концентрирование мочи.
Основным элементарным актом в концентрирующей системе почки является создание поперечного (горизонтального) осмотического градиента между восходящим и нисходящим коленами петли Генле. В результате их взаимодействия, и за счет противоточного перемещения канальцевой жидкости и крови, устанавливается определенная величина продольного (вертикального) осмотического градиента.
Но это одна часть концентрирующего механизма, теперь рассмотрим вторую - осмолярность окончательной мочи будет зависеть от проницаемости стенок собирательных трубок для воды, основным регулятором которой является АДГ. Если АДГ много, проницаемость стенок собирательной трубки для воды увеличивается и вода, подчиняясь осмотическому градиенту, существующему в интерстиции мозгового вещества, реабсорбируется и поступает в интерстиций и далее в кровь. Жидкость в собирательных трубках приходит в состояние равновесия с окружающим гиперосмолярным интерстицием, выделяется концентрированная моча. Если АДГ мало, стенки собирательных трубок становятся водонепроницаемыми, образуется гипотоническая моча, диурез увеличивается.
Запомните: Окончательный объём и состав мочи обусловлен функцией СТ. Их роль в осмотическом концентрировании и разведении мочи определяется как особенностями их анатомического расположения в почке, так и воздействием АДГ на проницаемость их стенок для воды. Другими словами, то, что произойдет с мочой в собирательной трубке и определит окончательное ее концентрирование.
Прежде чем перейти к механизму действия АДГ, обратим ваше внимание на то обстоятельство, что в отличие от проксимального канальца, где Nа и вода реабсорбировались вместе, в дистальном канальце и собирательной трубке вода и Nа реабсорбируются независимо. Именно это обстоятельство и позволяет дистальному отделу нефрона производить как концентрированную, так и разбавленную мочу. Дистальная реабсорбция называется факультативной (необязательной).
1. В процессе осмотического концентрирования мочи принимают участие петля Генле, дистальный каналец, собирательная трубка, сосуды и интерстиций мозгового вещества. Их оъединение в единый концентрирующий аппарат почки обусловлено их взаимным расположением и общностью протекающих в них процессов.
2. Процесс окончательного концентрирования мочи происходит в собирательной трубке за счет факультативной реабсорбции воды.
3. Условия для нее создает корково -сосочковый осмотический градиент мозгового вещества , созданный поворотно-противоточно-множительной системой петли Генле.
4. Факультативная реабсорбция воды в собирательной трубке регулируетсяАДГ.
5. В дистальном сегменте нефрона натрий и вода реабсорбируются независимо.
Петля Генле. Нисходящий ( тонкий ) отдел петли Генле. Дистальный отдел канальцев ( дистальные канальцы ). Собирательные трубки.
Сосудистый клубочек почки имеет 50 капиллярных петель, стенка которых является клубочковым фильтром. При электронно-микроскопическом исследовании нами установлено, что клубочковый фильтр состоит из трех элементов: эпителия клубочка, эндотелия его капилляров и располагающейся между ними базальной мембраны.
Эпителий клубочка (подоцит) состоит из крупного клеточного тела с ядром, митохондриями, аппаратом Гольджи и другими включениями. Цитоплазма имеет большие отростки - трабекулы, от которых в виде узких стебельков отходят малые отростки, или педикулы, которые своим внешним утолщенным концом «опираются» на базальную мембрану. Таким образом, трабекулы не лежат непосредственно на базальной мембране, а приподняты над ней благодаря педикулам, в результате чего создается так называемое подподоцитарное пространство. В этом пространстве переплетаются педикулы соседних клеток, образуя сложный лабиринт из щелей шириной от 300 до 1000 Å. Через которые фильтрат плазмы может поступать в полость капсулы Боумена - Шумлянского, минуя цитоплазму подоцита.
Эндотелий лежит на базальной мембране и пронизанной через довольно правильные промежутки порами размером от 600 до 1000 Å. Базальная мембрана клубочка - это непрерывный барьер между кровью и полостью капсулы Боумена - Шумлянского, который состоит по существу из двух мембран: наружной, субэпителиальной, которая продолжается в базальную мембрану капсулы Боумена - Шумлянского, и внутренней, субэндотелиальной, являющейся продолжением базальной мембраны артериол клубочка. Субэпителиальная мембрана довольно толстая, субэндотелиальная - тоньше и нежнее. Высокое содержание во фракции базальной мембраны коллагенового белка свидетельствует о тесной связи базальной мембраны гломерулярных капилляров с соединительной тканью клубочка, мезангиальные клетки которой способны нарабатывать вещество мембраны.
Между субэндотелиальной и субэпителиальной мембраной существует соединительная ткань, названная мезангиумом, который связывает капиллярные петли клубочка друг с другом и подвешивает их наподобие брыжейки к гломерулярному полюсу. Вместе с базальными мембранами он образует поддерживающий стержень клубочка. Мезангиум рассматривают как перикапиллярное или межкапиллярное интерстициальное пространство в котором найдены волокнистые структуры и интеркапиллярных, или мезангиальных клетки.
Изменения базальной мембраны клубочка характеризуются ее утолщением и гомогенизацией, возникновением пластинчатости, разрыхлением и фибриллярностью, а также появлением различных отложений.
Капсула клубочка состоит из эпителия и базальной мембраны, которая продолжается в проксимальный сегмент и по сравнению с базальной мембраной клубочка имеет иное строение. В ней различают внутренний - гомогенный и наружный - ретикулярный слой.
Эпителий капсулы представлен уплощенными клетками с четкими границами и выбухающим ядром. Они имеют немного митохондрий и не обладают той сложной организацией цитоплазмы, которая характерна для эпителия проксимальных канальцев.
Эпителиальные клетки капсулы отличаются весьма умеренной ферментативной активностью дегидрогеназ и диафораз. Исключение составляет дегидрогеназа изолимонной кислоты, содержание которой здесь довольно высоко. Отмечается также умеренная активность в этих клетках глюкозо-6-фосфатазы.
Эпителий капсулы крайне изменчив и в патологических условиях склонен к гиперпластическим реакциям. Кроме того, доказано, что изменчивость его структуры определяется рядом гормональных влияний.
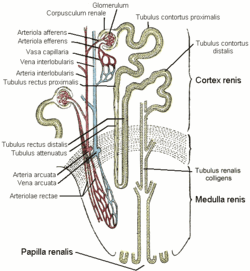
Канальцевую часть нефрона принято делить на три сегмента: проксимальный каналец, состоящий из извитой и прямой части (толстый нисходящий отдел петли Генле), тонкий сегмент петли Генле, дистальный каналец, состоящий из прямой части (толстый восходящий отдел петли Генле) и извитой части. Дистальные извитые канальцы впадают в систему собирательных трубочек.
Кровеносная система почки В почках имеются два функционально различных круга кровообращения: большой - кортикальный и малый - юкстамедуллярный. Кортикальный (большой) круг представлен сосудами коркового вещества: междольковыми артериями, приносящими сосудами и «чудесной сетью» основной массы гломерул, выносящими сосудами и постгломерулярной капиллярной и венозной сетью коры. Юкстамедуллярный (малый) круг кровообращения почки составляют: проксимальные отрезки интерлобулярных артерий, приносящие артериолы юкстамедуллярных клубочков и их сосудистая сеть, выносящие артериолы юкстамедуллярных клубочков, артериальные и венозные компоненты прямых сосудов пирамид, а также проксимальные отрезки интер лобулярных вен. Элементами юкстамедуллярного пути почечного кровотока могут быть межканальцевые капилляры кортикомедуллярной зоны, перигломерулярные сосудистые сплетения, связующие афферентные и эфферентные артериолы юкстамедуллярных клубочков, а также негломерулярные артериовенозные анастомозы пограничного слоя почки.
Более детально изменения представленных структур мы изучали на примере гломерулонефрита. При заболевании выявлено накопление материала, идентичного по составу базальной мембране, между пролиферирующими эндотелиальными и интеркапиллярными клетками и появление преципитата комплекса антиген-антитело чаще на эпителиальной стороне мембраны.
Гломерулонефриты являются самостоятельными нозологическими формами, но могут встречаться и при многих системных заболеваниях: системной красной волчанке, геморрагическом васкулите, подостром бактериальном эндокардите и др.
Прогрессирование гломерулонефрита после воздействия различных начальных факторов определяется достаточно стереотипными процессами, постепенно ведущими к склерозу. Это пролиферация мезангиальных клеток и накопление мезангиального матрикса, а также поражение стенки капилляров клубочка. В индукции гломерулонефрита ведущее место отводится иммунным факторам повреждения, реакциям гуморального или клеточного иммунитета. В дальнейшем прогрессировании важную роль приобретают медиаторы воспаления и неиммунные механизмы - гемодинамические и метаболические. На всех этапах развития ведущую роль играет персистирование этиологического фактора, известного, к сожалению, лишь у 1/10 больных.
а) Хронический гломерулонефрит с минимальными поражениями. При эллектронной. микроскопии выявляются изменения в базальной мембране, в основном в подоцитах. Выросты подоцитов сливаются, представляя собой единую систему с иммунными отложениями.
б) Мембранозный вариант нефрита: базальная мембрана поражается более глубоко. При обычной микроскопии утолщается, разрывы мембраны, большое отложение иммуноглобулинов и комплемента.
в) Фибропластический вариант: быстро идут процессы на базальной мембране, происходит активация мезангия, дальнейшее разрастание мембраны, разрастание клубочков идет медленнее.
г) Пролиферативный вариант: идет массовая пролиферация в основном сосудистого эндотелия (эндокапиллярный тип), реже пролиферация носит экстракапиллярный тип - со стороны Боумановской капсулы.
д) Мембранозно-пролиферативный. Специально выделяется гипертоническая форма хронического гломерулонефрита с клиникой классической формы гипертонической болезни, но есть изменения со стороны мочи.
Этиология гломерулонефритов до конца не выяснена. Известно только, что в их этиологии играют роль инфекции (наиболее четко при остром постстрептококковом гломерулонефрите), токсические вещества (органические растворители, алкоголь, ртуть, свинец и др.), экзогенные антигены, действующие с вовлечением иммунных механизмов, в том числе и в рамках гиперчувствительности немедленного типа (атопии), редко - эндогенные антигены - ДНК, мочевая кислота, опухолевые. Этиологический фактор удается установить у 80-70% больных острым гломерулонефритом и у 5-10% больных хроническим гломерулонефритом. У остальных больных причина болезни остается неизвестной.
В подавляющем большинстве случаев гломерулонефрит развивается при участии иммунных механизмов. При остром пострептококковом гломерулонефрите - это образование антител через 10-12 дней после фарингита или ангины, при хроническом гломерулонефрите - это чаще медленное образование иммунных комплексов, содержащих антиген и антитела к нему, откладывающихся в клубочках при недостаточности механизмов их удаления, у большинства больных быстропрогрессирующим гломерулонефритом - это образование антител к ткани клубочков (базальной мембране капилляров). Иммунные комплексы и антитела к базальной мембране могут быть выявлены и идентифицированы в ткани почек при иммуногистохимическом исследовании. Почечный клубочек отвечает на иммунное повреждение двумя видами патологических реакций: пролиферацией клеток почечного клубочка (мезангиальных, эндотелиальных, эпителиальных) и выработкой этими клетками межклеточного вещества. Оба процесса стимулируют цитокины (в первую очередь интерлейкин-1, фактор некроза опухолей, тромбоцитарный фактор роста, трансформирующй фактор роста бета), которые выделяются клетками, инфильтрирующими почечный клубочек.
Таким образом, гломерулонефрит - это двухстороннее воспалительное заболевание почек иммунного генеза, которое характеризуется постепенной, но неуклонной гибелью клубочков, сморщиванием почки, понижением функции, развитием артериальной.
Петля Генле
Нефрон (от греческого νεφρός (нефрос) — «почка») — структурно-функциональная единица почки животного. Нефрон состоит из мальпигиева тельца, где происходит фильтрация, и системы канальцев, в которых осуществляются реабсорбция (обратное всасывание) и секреция веществ. Нефрон регулирует кровяное давление, контролирует уровень электролитов и метаболитов, а также принимает участие в поддержании постоянного уровня pH. Нефрон также является железой внутренней секреции, осуществляя синтез таких стероидных гормонов, как антидиуретический гормон, альдостерон и паратиреоидный гормон.
Содержание
Структура и функция
Почечное тельце
Нефрон начинается с почечного тельца, которое состоит из клубочка и капсулы Боумена-Шумлянского. Здесь осуществляется ультрафильтрация плазмы крови, которая приводит к образованию первичной мочи.
Клубочек
Клубочек представляет собой группу сильно фенестрированных (окончатых) капилляров, получающих кровоснабжение от афферентной артериолы. Гидростатическое давление крови создаёт движущую силу для фильтрации жидкости и растворённых веществ в просвет капсулы Боумена-Шумлянского. Непрофильтровавшаяся часть крови из клубочков поступает в эфферентную артериолу. Эфферентная артериола поверхностно расположенных клубочков распадается на вторичную сеть капилляров, оплетающих извитые канальцы почек, эфферентные артериолы от глубоко расположенных (юкстамедуллярных) нефронов продолжаются в нисходящие прямые сосуды (vasa recta), опускающиеся в мозговое вещество почек. Вещества, реабсорбированные в канальцах, в дальнейшем поступают в эти капиллярные сосуды.
Капсула Боумена-Шумлянского
Капсула Боумена-Шумлянского окружает клубочек и состоит из висцерального (внутреннего) и париетального (внешнего) листков. Внешний листок представляет собой обычный однослойный плоский эпителий. Внутренний листок составлен из подоцитов, которые лежат на базальной мембране эндотелия капилляров, и ножки которых покрывают поверхность капилляров клубочка. Ножки соседних подоцитов образуют на поверхности капилляра интердигиталии. Промежутки между клетками в этих интердигиталиях и образуют, собственно, щели фильтра, затянутые мембраной. Размер этих фильтрационных пор ограничивает перенос крупных молекул и клеточных элементов крови.
Между внутренним листком капсулы и внешним, представленным простым, непроницаемым, плоским эпителием, лежит пространство, в которое поступает жидкость, профильтровавшаяся через фильтр, который сформирован мембраной щелей в интердигиталиях, базальной пластинкой капилляров и гликокаликсом, секретируемым подоцитами.
Нормальная скорость клубочковой фильтрации (СКФ) составляет 180—200 литров в сутки, что в 15-20 раз превышает объём циркулирующей крови — иными словами, вся жидкость крови за сутки успевает профлильтроваться приблизительно двадцать раз. Измерение СКФ является важной диагностической процедурой, её снижение может быть показателем почечной недостаточности.
Небольшие молекулы — такие, как вода, ионы Na + , Cl - , аминокислоты, глюкоза, мочевина, одинаково свободно проходят через клубочковый фильтр, так же проходят через него белки массой до 30 Кд, хотя, поскольку белки в растворе обычно несут отрицательный заряд, для них определённое препятствие составляет отрицательно заряженный гликокаликс. Для клеток и более крупных белков клубочковый ультрафильтр представляет непреодолимое препятствие. В результате, в пространство Шумлянского-Боумена, и далее в проксимальный извитой каналец, поступает жидкость, по составу отличающаяся от плазмы крови только отсутствием крупных белковых молекул.
2. Строение нефрона. Механизм образования мочи
В корковом слое почки находятся почечные капсулы (капсулы нефрона), внутри каждой из которых располагается капиллярный клубочек.
В мозговом (пирамидальном) слое находятся извитые канальцы. Канальцы собираются вместе в собирательные трубочки, впадающие в почечную лоханку. От почечной лоханки каждой почки отходит мочеточник, соединяющий почку с мочевым пузырём.
От капсулы отходит извитой каналец первого порядка (проксимальный извитой каналец). Он выходит в мозговой слой и образует петлю Генле. Петля Генле переходит в извитой каналец второго порядка (дистальный извитой каналец), а тот впадает в собирательную трубочку, ведущую к лоханке.
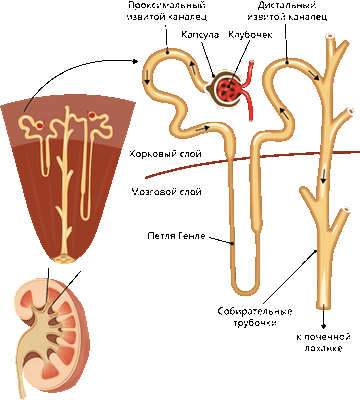
Почечная артерия разделяется на приносящие артериолы. Каждая артериола ветвится и образует капиллярный клубочек капсулы нефрона.
На выходе из капсулы капилляры сливаются в выносящую артериолу, которая разветвляется на вторичную сеть капилляров, оплетающую извитые канальцы и петлю Генле.
Из капилляров кровь поступает в венулы, сливающиеся в почечную вену, и течёт по ней к нижней полой вене.
Моча образуется в почках из крови, которой почки хорошо снабжаются. Мочеобразование состоит из двух процессов — фильтрации и реабсорбции.
Сначала кровь, поступающая в капиллярный клубочек по приносящей артериоле, фильтруется через стенки капилляров в полость капсулы нефрона.
В капиллярах клубочков давление крови высокое. Поэтому вода и молекулы растворённых в плазме веществ фильтруются сквозь тонкие стенки капилляров и поступают в почечный каналец. Образовавшийся фильтрат называют первичной мочой. По составу она похожа на плазму крови, но не содержит белков. В состав первичной мочи входят как продукты обмена (мочевина и мочевая кислота), так и необходимые организму вещества (глюкоза, аминокислоты, витамины и т. д.).
В извитых канальцах происходит реабсорбция, т. е. обратное всасывание в кровь первичной мочи и образование вторичной (конечной) мочи. Возвращается в кровь большая часть воды, а также аминокислоты, глюкоза, витамины, некоторые соли.
Во вторичной моче остаётся мочевина и мочевая кислота. Их содержание возрастает в десятки раз. Намного больше в ней также ионов калия, а содержание ионов натрия остаётся тем же.
За сутки образуется около \(150\) л первичной мочи и около \(1,5\) л в сутки вторичной мочи, что составляет примерно \(1\) % объёма первичной мочи. С первичной мочой удаляются из организма ненужные вещества, а все полезные вещества поступают обратно в кровь.
Вторичная моча из канальцев попадает в почечную лоханку, а затем по мочеточникам стекает в мочевой пузырь и по мочеиспускательному каналу выводится наружу.
Нервная регуляция осуществляется гипоталамусом, к которому по нейронам автономной нервной системы поступают сигналы о составе и давлении крови от рецепторов, расположенных в стенках кровеносных сосудах.
Гуморальная регуляция происходит с участием гормонов разных желёз: гипофиза, коры надпочечников, паращитовидных.
Петля Генле
Сущность работы поворотно-противоточной cистемы состоит в том, что два колена петли — нисходящее и восходящее, тесно соприкасаясь друг с другом, функционируют сопряженно как единый механизм ( рис. 103 ). Эпителий нисходящего (проксимального) отдела петли пропускает воду, но не пропускает ионы натрия. Эпителий же восходящего (дистального) отдела петли обладает способностью активно реабсорбировать ионы натрия, т. е. переводить их из канальцевой мочи в тканевую жидкость почки, но одновременно он не пропускает воду из просвета канальцев в тканевую жидкость.
При прохождении мочи через нисходящие (проксимальные) отделы петли Генле моча постепенно сгущается вследствие перехода воды в тканевую жидкость. Переход воды — пассивный процесс, обусловленный тем, что эпителий расположенного рядом восходящего (дистального) отдела петли активно реабсорбирует ионы натрия, т. е. переводит их из просвета канальца в тканевую (интерстициальную) жидкость; перешедшие в тканевую жидкость ионы натрия притягивают туда молекулы воды, но не из дистального, а из проксимального канальца.
Рис. 103. Схема работы поворотно-противоточной системы (по Бесту и Тейлору). Затемненный фон показывает величину концентрации мочи и тканевой жидкости. Белые стрелки — выделение воды, черные стрелки — выделение ионов натрия. 1 — извитой каналец первого порядка, переходящий в проксимальный отдел петли; 2 — извитой каналец второго порядка, выходящий из дистального отдела петли; 3 — собирательная трубка.
Выход воды из просвета проксимального канальца приводит к тому, что моча по ходу этого канальца сгущается все больше и больше в области вершины петли становится наиболее концентрированной. Высокая концентрация мочи способствует переходу ионов натрия из мочи дистального отдела петли в тканевую жидкость, поскольку стенки дистального канальца непроницаемы для воды, но активно реабсорбируют ионы натрия. Переход ионов натрия из дистального канальца петли в тканевую жидкость в свою очередь повышает ее осмотическое давление, что, как указывалось выше, обусловливает выход воды из проксимального канальца.
Таким образом, выход воды из мочи в тканевую жидкость в проксимальном канальце способствует реабсорбции натрия в дистальном канальце, а реабсорбция натрия в свою очередь обусловливает выход воды из проксимального канальца. Оба эти процесса сопряжены. Вследствие выхода натрия из мочи в тканевую жидкость гипертоничная у вершины петли моча становится затем изотоничной или даже гипотоничной (по отношению к плазме крови) в конце восходящего канальца петли Генле .
Разница между осмотическим давлением мочи в тесно прилегающих друг к другу нисходящем и восходящем канальцах на каждом данном уровне петли невелика. Осмотическое давление тканевой жидкости, окружающей канальцы, примерно таково же, как и осмотическое давление мочи в данном участке проксимального и дистального канальцев.
Осмотическое давление мочи в нисходящем канальце вследствие всасывания воды постепенно повышается, а осмотическое давление мочи в восходящем канальце вследствие реабсорбции натрия столь же постепенно понижается. Таким образом, между двумя соседними участками проксимаьного (или дистального) канальца разность осмотического давления невелика. По ходу же петли эти небольшие перепады давления в каждом из участков канальца суммируются, что приводит к очень большому перепаду (градиненту) осмотического давления между началом (или концом) петли и ее вершиной. Необходимо подчеркнуть, что в начало петли поступает значительное количество мочи, нзотоничной крови. В петле Генле моча теряет большое количество воды и натрия и из петли вытекает значительно меньшее количество мочи, вновь изотоничной или даже гипотоничной по отношению к плазме крови. Петля, таким образом, работает как концентрационный механизм, приводящий к реабсорбции большого количества воды и ионов натрия. Принцип поворотно-противоточной системы, аналогичный описанному выше, применяют в технике, когда необходимо создать большие разности концентрации каких-либо веществ.
В извитых канальцах второго порядка происходит дальнейшее всасывание ионов натрия, калия, воды и других веществ. В отличие от процессов, протекающих в извитых канальцах первого порядка и в петле Генгле , величина реабсорбции ионов натрия в калия в извитых канальцах первого порядка является не постоянной (обязательная реабсорбция), а изменчивой (факультативная реабсорбция). Она зависит от уровня ионов натрия и калия в крови и является важным регуляторным механизмом, поддерживающим постоянство концентрации этих ионов в организме.
Читайте также:
- Показания для эндоскопического релиза кубитального канала
- Оболочки спинного и головного мозга. Виды оболочек спинного и головного мозга.
- Нарушения миграции почек
- Введение в лучевую диагностику лимфатических узлов шеи: лучевая анатомия, методы исследования
- Последовательность исследования средостения. Дополнительная оценка средостения