Регуляция дыхания. Регуляция вентиляции легких.
Добавил пользователь Morpheus Обновлено: 21.01.2026
Общая характеристика процесса дыхания. Дыхание - это совокупность процессов, обеспечивающих обмен газов между организмом и окружающей средой. В результате дыхания происходит потребление организмом кислорода и выделение углекислого газа. Процесс дыхания состоит из трех основных этапов: обмен газов в легких между организмом и средой (внешнее или легочное дыхание); перенос газов кровью (кислорода из легких к тканям и углекислого газа от тканей к легким); газообмен между кровью и тканями (внутреннее или тканевое дыхание).
Механизм вдоха и выдоха. При вдохе происходит расширение грудной полости в результате сокращения наружных межреберных мышц и диафрагмы. Так как давление в плевральной полости отрицательное, при расширении грудной полости расширяются и легкие. Давление внутри легких становится ниже атмосферного, и наружный воздух проходит в легкие. При усиленном дыхании в акте вдоха участвуют все мышцы, способные поднимать ребра и грудину: большие и малые грудные, лестничные, грудинно-ключично-сосцевидные мышцы плечевого пояса. Выдох наступает в результате уменьшения объема грудной полости при расслаблении наружных межреберных мышц. При активном выдохе сокращаются и мышцы брюшной стенки (косые, поперечные и прямые), что усиливает поднятие диафрагмы. У человека в состоянии покоя цикл дыхания, состоящий из вдоха и выдоха, повторяется 16-20 раз в минуту.
Объемы легочного воздуха. Человек в состоянии покоя вдыхает и выдыхает около 500 мл воздуха. Этот объем называют дыхательным объемом (ДО). Если после спокойного вдоха сделать усиленный дополнительный вдох, то в легкие может поступить еще 1500 мл воздуха. Этот объем называют резервным объемом вдоха (РОВД). После спокойного выдоха можно при максимальном напряжении дыхательных мышц выдохнуть еще 1500 мл воздуха. Этот объем носит название резервного объема выдоха (РОвыд.). В сумме дыхательный объем, резервный объем вдоха и резервный объем выдоха составляют жизненную емкость легких (ЖЕЛ). Нормальная ЖЕЛ составляет в среднем у женщин 2700 мл, а у мужчин 3500 мл.
Даже после максимального выдоха в легких еще остается около 1200 мл воздуха, который носит название остаточного объема (ОО). Объемы легочного воздуха и ЖЕЛ измеряют при помощи спирометра и спирографа.
Вентиляция легких.Вентиляцией легких называют объем воздуха, проходящий через легкие в 1 мин. Иначе его называют минутным объемом дыхания (МОД). В покое МОД равен 5-8 л/мин, при мышечной работе он увеличивается и нередко достигает 80-150 л/мин. Человек вдыхает атмосферный воздух, в котором содержится 20,94% кислорода, 79,03% азота и незначительное количество углекислого газа - 0,03%. Выдыхаемый воздух содержит кислород (16,3%), азот (79,7%) и много углекислого газа (4%). Процентное содержание газов в воздухе определяют при помощи газоанализатора.
Обмен газов в легких. Перенос кислорода из альвеолярного воздуха в кровь и углекислого газа из крови в альвеолярный воздух происходит путем диффузии. Ее движущей силой служат разности (градиенты) парциальных давлений (напряжений) кислорода и углекислого газа по обе стороны аэрогематического барьера. Никакого механизма активного транспорта газов здесь не существует. Парциальное давление кислорода в альвеолярном воздухе равно 102 мм рт. ст., а парциальное давление углекислого газа - 40 мм рт. ст. В притекающей к капиллярам легких венозной крови давление кислорода составляет 40, а углекислого газа - 47 мм рт.ст. Поскольку парциальное давление кислорода в альвеолах больше, чем в венозной крови, то он диффундирует из альвеол в капилляры. Напротив, напряжение углекислого газа больше в венозной кроки, чем в альвеолярном воздухе, поэтому он диффундирует в альвеолы.
Обмен газов в тканях. В легких венозная кровь превращается в
артериальную. Артериальная кровь направляется к тканям. Газообмен в
тканях происходит по тому же принципу. В артериальной крови напряжение
кислорода равно 100 мм рт. ст., а углекислого газа - 40 мм рт. ст. В
тканях напряжение кислорода близко к нулю, а напряжение углекислого
газа составляет около 60 мм рт. ст. Вследствие разности давления
углекислый газ из тканей диффундирует в кровь, а кислород - в ткани. Кровь становится венозной и по венам поступает в легкие, где цикл обмена
газов повторяется.
Транспорт газов кровью. Кислород в основном транспортируется к тканям в составе оксигемоглобина. Лишь небольшая часть кислорода (около 2%), переносимого кровью, растворена в плазме. Плазмой крови в физически растворенном состоянии переносится 3-6% углекислого гaза. В эритроцитах примерно 10% углекислого газа соединяется с гемоглобином и образуется карбгемоглобин. Остальная часть соединяется с водой и превращается в угольную кислоту.
Эта реакция катализируется ферментом карбоангидразой. В тканевых капиллярах угольная кислота реагирует с ионами Na + , K + и превращается в бикарбонаты.
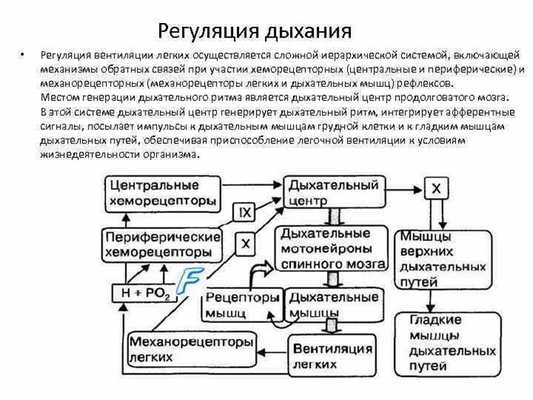
Регуляция дыхания. Дыхание регулируется дыхательным
центром, который находится в продолговатом мозге. Дыхательный центр
состоит из центров вдоха и выдоха. Кроме того, нейроны, связанные с
регуляцией дыхания, находятся в варолиевом мосту. Здесь выделяют
пневмотаксический центр, который участвует в переключении фаз
дыхательного цикла.
Характерная особенность дыхательного центра - автоматия. Автоматия обуславливает ритмичность дыхания. На работу дыхательного центра оказывает влияние кора больших полушарий головного мозга. Человек произвольно регулирует дыхание при разговоре, пении; он может задерживать дыхание.
Особое значение в регуляции дыхания имеют рефлексы от хеморецепторов, чувствительных к изменению напряжения в крови углекислого газа и кислорода. Эта сигнализация исходит от центральных (бульбарных) и периферических (артериальных) хеморецепторов. Нервные и гуморальные механизмы регуляции дыхания тесно взаимосвязаны.
Модуль № 1. Физиология дыхания
Дыхательный центр продолговатого мозга, его функции.
Ритмическая последовательность вдоха и выдоха, а также изменение характера дыхательных движений в зависимости от состояния организма (покой, работа различной интенсивности, эмоциональные проявления и т. д.) регулируются дыхательным центром, расположенным в продолговатом мозге. Дыхательным центром называется совокупность нейронов, обеспечивающих деятельность аппарата дыхания и его приспособление к изменяющимся условиям внешней и внутренней среды.
В дыхательном центре имеются две группы нейронов: инспираторные и экспираторные. Обнаружены некоторые особенности в работе дыхательного центра. При спокойном дыхании активна только небольшая часть дыхательных нейронов, и, следовательно, в дыхательном центре есть резерв нейронов, который используется при повышенной потребности организма в кислороде. Установлено, что между инспираторными и экспираторными нейронами дыхательного центра существуют функциональные взаимосвязи. Они выражаются в том, что при возбуждении инспираторных нейронов, обеспечивающих вдох, деятельность экспираторных нервных клеток заторможена, и наоборот. Таким образом, одной из причин ритмичной, автоматической деятельности дыхательного центра являются взаимосвязанные функциональные отношения между этими группами нейронов.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III—IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах (III—XII) грудных сегментов спинного мозга.
Регуляция деятельности дыхательного центра
Механизм первого вдоха новорожденного. В организме матери газообмен плода происходит через пупочные сосуды, тесно контактирующие с плацентарной кровью матери. После рождения ребенка и отделения плаценты указанная связь нарушается. Метаболические процессы в организме новорожденного приводят к образованию и накоплению углекислого газа, который, так же как и недостаток кислорода, гуморально возбуждает дыхательный центр. Кроме того, изменение условий существования ребенка приводит к возбуждению экстеро- и проприорецепторов, что также является одним из механизмов, принимающих участие в осуществлении первого вдоха новорожденного.
Влияние клеток коры большого мозга на активность дыхательного центра.
Учащение и углубление дыхания, которые наступают до начала физической работы или спортивных состязаний, осуществляются по механизму условных рефлексов. Эти изменения в дыхательных движениях отражают сдвиги в активности дыхательного центра и имеют приспособительное значение, способствуя подготовке организма к выполнению работы, требующей большой затраты энергии и усиления окислительных процессов.
По мнению М.Е. Маршака, корковая регуляция дыхания обеспечивает необходимый уровень легочной вентиляции, темп и ритм дыхания, постоянство уровня углекислого газа в альвеолярном воздухе и артериальной крови.
По М.В. Сергиевскому, регуляция активности дыхательного центра представлена тремя уровнями.
Первый уровень регуляции — спинной мозг. Здесь располагаются центры диафрагмальных и межреберных нервов, обусловливающие сокращение дыхательных мышц.
Второй уровень регуляции — продолговатый мозг. Здесь находится дыхательный центр, который перерабатывает разнообразные афферентные импульсы, идущие от дыхательного аппарата, а также от основных рефлексогенных сосудистых зон
Третий уровень регуляции — верхние отделы головного мозга, включающие и корковые нейроны. Только при участии коры большого мозга возможно адекватное приспособление реакций системы дыхания к изменяющимся условиям окружающей среды.
Лекция 20 Регуляция дыхания
Регуляция дыхания осуществляется путем рефлекторных реакций, возникающих в результате возбуждения специфических рецепторов, заложенных в легочной ткани, сосудистых рефлексогенных зонах и других участках. Центральный аппарат регуляции дыхания представляют образования спинного мозга, продолговатого мозга и вышележащих отделов нервной системы. Основная функция управления дыханием осуществлянется дыхательными нейронами ствола головного мозга, которые передают ритмические сигналы в спинной мозг к мотонейронам дыхательных мышц.
Дыхательный нервный центр - это совокупность нейронов центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и постоянное приспособление внешнего дыхания к изменяющимся условиям внутри организма и в окружающей среде. Основная (рабочая) часть дыхательного нервного центра расположена в продолговатом мозгу. В ней различают два отдела: инспираторный (центр вдоха) и экспираторный (центр выдоха). Дорсальная группа дыхательных нейронов продолговатого мозга состоит преимущественно из инспираторных нейронов. Они частично дают поток нисходящих путей, вступающих в контакт с мотонейронами диафрагмального нерва. Вентральная группа дыхательных нейронов посылает преимущественно нисходящие волокна к мотонейронам межреберных мышц. В передней части варолиева моста обнаружена область, названная пневмотаксическим центром. Этот центр имеет отношение к работе как экспи-, так и инспираторного его отделов. Важной частью дыхательного нервного центра является группа нейронов шейного отдела спинного мозхга (III-IV шейные сегменты), где расположены ядра диафрагмальных нервов.
К моменту рождения ребенка дыхательный центр способен давать ритмическую смену фаз дыхательного цикла, но эта реакция очень несовершенна. Дело заключается в том, что к рождению дыхательный центр еще не сформирован, его формирование заканчивается к 5-6 годам жизни. Это подтверждается тем, что именно к этому периоду жизни детей дыхание у них становится ритмичным и равномерным. У новорожденных же оно неустойчиво как по частоте, так и глубине и ритму. У них дыхание диафрагмальное и практически мало отличается во время сна и бодроствования (частота от 30 до 100 в минуту). У детей 1 года количество дыхательных движений днем в пределах 50-60, а ночью - 35-40 в минуту, неустойчивое и диафрагмальное. В возрасте 2-4 лет - частота становится в пределах 25-35 и носит преимущественно диафрагмальный тип. У 4-6 - летних детей частота дыхания 20-25, смешанное - грудное и диафрагмальное. К 7 -14 годам достигает уровня 19-20 в минуту, оно является в это время смешанным. Таким образом, окончательное формирование нервного центра практически относится к этому возрастному периоду.
Как же происходит возбуждение дыхательного центра? Один из важнейших путей его возбуждения - это автоматия. Единой точки зрения на природу автоматии нет, но имеются данные о том, что в нервных клетках дыхательного центра возможно возникновение вторичной деполяризации (по принципу диастолической деполяризации в сердечной мышце), которая, достигая критического уровня, и дает новый импульс. Однако одним из основных путей возбуждения дыхательного нервного центра является его раздражение углекислотой. На прошлой лекции мы отметили, что углекислоты много остается в крови, оттекающей от легких. Она и выполняет функцию основого раздражителя нервных клеток продолговатого мозга. Это опосредуется через специальные образования - хеморецепторы, расположенные непосредственно в структурах продолговатого мозга («центральные хеморецепторы»). Они очень чувствительны к напряжению углекислого газа и кислотно-щелочному состоянию омывающей их межклеточной мозговой жидкости.
Углекислота может легко диффундировать из кровеносных сосудов головного мозга в спинномозговую жидкость и стимулировать хеморецепторы продолговатого мозга. Это еще один путь возбуждения дыхательного центра.
Наконец, его возбуждение может осуществляться и рефлекторно. Все рефлексы, обеспечивающие регуляцию дыхания мы условно подразделяем на: собственные и сопряженные.
Рецепторы растяжения легких находятся, преимущественно в гладких мышцах воздухоносных путей (трахее, бронхах). Таких рецепторов в каждом легком около 1000 и связаны они с дыхательным центром крупными миелинизированными афферентными волокнами блуждающего нерва с высокой скоростью проведения. Непосредственным раздражителем этого типа механорецепторов является внутреннее напряжение в тканях стенок воздухоносных путей. При растяжении легких во время вдоха частота этих импульсов возрастает. Раздувание легких вызывает рефлекторное торможение вдоха и переход к выдоху. При перерезке блуждающих нервов эти реакции прекращаются, и дыхание становится замедленным и глубоким. Указанные реакции называют рефлексом Геринга-Брейера. Этот рефлекс воспроизводится у взрослого человека, когда дыхательный объем превосходит 1 л (при физической нагрузке, например). Он имеет большое значение у новорожденных.
Ирритантные рецепторы или быстро адаптирующиеся механорецепторы воздухоносных путей, рецепторы слизистой оболочки трахеи и бронхов. Они реагируют на резкие изменения объема легких, а также при действии на слизистую трахеи и бронхов механических или химических раздражителей (пылевых частиц, слизи, паров едких веществ, табачного дыма и т.п.). В отличие от легочных рецепторов растяжения ирритантные рецепторы обладают быстрой адаптацией. При попадании в дыхательные пути мельчайших инородных тел (пыли, частиц дыма), активация ирритантных рецепторов вызывает у человека кашлевой рефлекс. Его рефлекторная дуга такова - от рецепторов информация через верхнегортанный, языкоглоточный, тройничный нерв идет к соотвествующим структурам мозга, отвечающим за выдох (срочный выдох - кашель). Если изолированно возбуждаются рецепторы носовых дыхательных путей, то это вызывает другой срочный выдох - чихание.
Юкстакапиллярные рецепторы - расположены вблизи капилляров альвеол и дыхательных бронхов. Раздражителем этих рецепторов является повышение давления в малом круге кровообращения, а также увеличение объема интерстициальной жидкости в легких. Это наблюдается при застое крови в малом круге кровообращения, отеке легких, повреждениях легочной ткани (например, при пневмонии). Импульсы от этих рецепторов направляются к дыхательному центру по блуждающему нерву, вызывая появление частого поверхностного дыхания. При заболеваниях вызывает ощущение одышки, затрудненного дыхания. Может быть не только учащенное дыхание (тахипное), но и рефлекторное сужение бронхов.
Еще различают большую группу собственных рефлексов, которые берут свое начало от проприорецепторов дыхательной мускулатуры. Рефлекс от проприорецепторов межреберных мышц осуществляется во время вдоха, когда эти мышцы, сокращаясь, посылают информацию через межреберные нервы к экспираторному отделу дыхательного центра и в результате наступает выдох. Рефлекс от проприорецепторов диафрагмы осуществляется в ответ на ее сокращение во время вдоха, в результате информация поступает по диафрагмальным нервам вначале в спинной, а потом в продолговатый мозг в экспираторный отдел дыхательного центра и наступает выдох.
Таким образом, все собственные рефлексы дыхательной системы осуществляются во время вдоха и заканчиваются выдохом.
Сопряженные рефлексы дыхательной системы - это рефлексы, которые начинаются за ее пределами. К этой группе рефлексов, прежде всего, относится рефлекс на сопряжение деятельности системы кровообращения и дыхания. Такой рефлекторный акт начинается от периферических хеморецепторов сосудистых рефлексогенных зон. Наиболее чувствительные из них находятся в области синокаротидной зоны. Синокаротидный хеморецептивный сопряженный рефлекс - осуществляется при накоплении углекислого газа в крови. Если его напряжение растет, то происходит возбуждение наиболее высоковозбудимых хеморецепторов (а они именно в этой зоне и находятся в синокаротидном тельце), возникающая волна возбуждения идет от них по IX паре черпномозговых нервов и достигает экспираторного отдела дыхательного центра. Возникает выдох, который и усиливает выброс лишней углекислоты в окружающее пространство. Таким образом, система кровообращения (она, кстати, при осуществлении этого рефлекторного акта также работает более интенсивно, возрастает частота сердечных сокращений, скорость кровотока) влияет на деятельность системы дыхания.
Другой разновидностью сопряженных рефлексов дыхательной системы является многочисленная группа экстероцептивных рефлексов. Они берут свое начало от тактильных (вспомните реакцию дыхания на осязание, прикосновение), температурных (тепло - увеличивает, холод - уменьшает дыхательную функцию), болевых (слабые и средней силы раздражители - усиливают, сильные - угнетают дыхание) рецепторов.
Проприорецептивные сопряженные рефлексы дыхательной системы осуществляются вследствие раздражения рецепторов скелетных мышц, суставов, связок. Это наблюдается при выполнении физической нагрузки. Почему это происходит? Если в состоянии покоя человеку необходимо 200-300 мл кислорода в минуту, то при физической нагрузке этот объем должен значительно возрости. В этих условиях увеличивается и МО, артериовенозная разница по кислороду. Увеличение этих показателей сопровождается повышением потребления кислорода. Далее все зависит от объема работы. Если работа длится 2-3 минуты и мощность ее достаточно велика, то потребление кислорода непрерывно растет с самого начала работы и снижается лишь после ее прекращения. Если же продолжительность работы больше, то потребление кислорода, нарастая в первые минуты, поддерживается в дальнейшем на постоянном уровне. Потребление кислорода возрастает тем более, чем тяжелее физическая работа. Наибольшее количество кислорода, которое организм может поглотить за 1 минуту при предельно тяжелой для него работе, называется максимальное потребление кислорода (МПК). Работа, при которой человек достигает своего уровня МПК, должна длиться не более 3 минут. Существует много способов определения МПК. У не занимающихся спортом или физическими упражнениями людей величина МПК не превышает 2,0-2,5 л/мин. У спортсменов она может быть выше более чем в два раза. МПК является показателем аэробной производительности организма. Эта способность человека совершать очень тяжелую физическую работу, обеспечивая свои энергетические расходы за счет кислорода, поглощаемого непосредственно во время работы. Известно, что даже хорошо тренированный человек может работать при потреблении кислорода на уровне 90-95% от уровня своего МПК не более 10-15 минут. Тот, кто имеет большую аэробную производительность, тот достигает лучших результатов в работе (спорте) при относительно одинаковой технической и тактической подготовленности.
Почему при физической работе возникает увеличение потребления кислорода? В этой реакции можно выделить несколько причин: раскрытие дополнительных капилляров и увеличение крови в них, сдвиг кривой диссоциации гемоглобина вправо и вниз, увеличение температуры в мышцах. Для того, чтобы мышцы могли совершать определенную работу, им нужна энергия, запасы которой в них восстанавливаются при доставке кислорода. Таким образом, существует зависимость между мощностью работы и количеством кислорода, которое требуется для работы. То количество крови, которое требуется для работы, называется кислородным запросом. Кислородный запрос может достигать при тяжелой работе до 15-20 л в минуту и более. Однако максимум потребления кислорода в два-три раза меньше. Можно ли выполнить работу, если минутный кислородный запас превышает МПК? Чтобы правильно ответить на этот вопрос, надо вспомнить, для чего используется кислород при мышечной работе. Он необходим для восстановления богатых энергией химических веществ, обеспечивающих мышечное сокращение. Кислород обычно взаимодействует с глюкозой, и она, окисляясь, освобождает энергию. Но глюкоза может расщепляться и без кислорода, т.е. анаэробным путем, при этом тоже выделяется энергия. Кроме глюкозы, есть и другие вещества, способные расщепляться без кислорода. Следовательно, работа мышц может быть обеспечена и при недостаточном поступлении кислорода в организм. Однако в этом солучае образуется много кислых продуктов и для их ликвидации нужен кислород, ибо они разрушаются путем окисления. То количество кислорода, которое требуется для окисления продуктов обмена, образовавшихся при физической работе, называется кислородный долг. Он возникает во время работы и ликвидируется в восстановительном периоде после нее. На его ликвидацию уходит от нескольких минут до полутора часов. Все зависит от длительности и интенсивности работы. Основную роль в образовании кислородного долга составляет молочная кислота. Чтобы продолжить работу при наличии в крови большого ее количества, организм должен иметь мощные буферные системы и его ткани должны быть приспособлены к работе при недостатке кислорода. Такое приспособление тканей служит одним из факторов, обеспечивающих высокую анаэробную производительность.
Все это усложняет регуляцию дыхания при физической работе, так как потребление кислорода в организме возрастает и его недостаток в крови приводит к раздражению хеморецепторов. Сигналы от них идут в дыхательный центр, в результате дыхание учащается. При мышечной работе много образуется углекислоты, которая поступает в кровь и она может действовать на дыхательный центр непосредственно черех центральные хеморецепторы. Если недостаток кислорода в крови приводит преимущественно к учащению дыхания, то избыток углекислоты вызывает его углубление. При физической работе оба эти фактора действуют олновременно, вследствие чего происходит и учащение, и углубление дыхания. Наконец, импульсы идущие от работающих мышц, достигают дыхательного центра и усиливают его работу.
При функционировании дыхательного центра все отделы его функционально взаимосвязаны. Это достигается следующим механизмом. При накоплении углекислоты возбуждается инспираторный отдел дыхательного центра, от него информация идет в пневматоксический отдел центра, потом к экспираторному его отделу. Последний, кроме того, возбуждается за счет целой гаммы рефлекторных актов (с рецепторов легких, диафрагмы, межреберных мышц, дыхательных путей, хеморецепторов сосудов). Вследствие его возбуждения через специальный тормозный ретикулярный нейрон угнетается деятельность центра вдоха и на смену ему приходит выдох. Так как центр вдоха тормозится, то он не посылает далее импульсы в пневматоксический отдел, а от него прекращается поток информации в центр выдоха. К этому моменту накапливается в крови углекислота и снимаются тормозные влияния со стороны экспираторного отдела дыхательного центра. Вследствие такого перераспределения потока информации возбуждается центр вдоха и наступает на смену выдоху вдох. И все вновь повторяется.
Важным элементом в регуляции дыхания является блуждающий нерв. Именно через его волокна идут основные влияния на центр выдоха. Поэтому в случае его повреждения (также как и при повреждении пневматоксического отдела дыхательного центра) дыхание изменяется так, что вдох остается нормальным, а выдох резко затягивается. Такой тип дыхания называют вагус-диспноэ.
Мы уже отмечали выше, что при подъме на высоту происходит увеличение легочной вентиляции, обусловленное стимуляцией хеморецепторов сосудистых зон. Одновременно с этим возрастает частота сердечных сокращений и МО. Эти реакции несколько улучшают кислородный транспорт в организме, но не надолго. Поэтому при длительном пребывании в горах по мере адаптации к хронической гипоксии начальные (срочные) реакции дыхания постепенно уступают место более экономному приспособлению газотранспортной системы организма. Так, у постоянных жителей больших высот реакция дыхания на гипоксию оказывается резко ослабленной (гипоксическая глухота) и легочная вентиляция поддерживается почти на том же уровне, что и у живущих на равнине. Зато при длительном проживании в условиях высокогорья возрастает ЖЕЛ, повышается КЕК, в мышцах становится больше миоглобина, в митохондриях усиливается активность ферментов, обеспечивающих биологическое окисление и гликолиз. У людей, живущих в горах, кроме того, понижена чувствительность тканей организма, в частности, центральной нервной системы, к недостаточному снабжению кислородом.
На высотах боле 12000 м давление воздуха очень мало и в этих условиях даже дыхание чистым кислородом не решает проблемы. Поэтому при полетах на этой высоте необходимы герметические кабины (самолеты, космические корабли).
Человеку иногда приходиться работать и в условиях повышенного давления (водолазные работы). На глубине в крови начинает растворяться азот и при быстром подъеме из глубины он не успевает выделяться из крови, газовые пузырьки вызывают эмболию сосудов. Состояние, возникающее при этом, называется кесонная болезнь. Она сопровождается болями в суставах, головокружением, одышкой, потерей сознания. Поэтому азот в смесях воздуха заменяют нерастворимыми газами (например, гелием).
Человек может произвольно задерживать дыхание не более чем на 1-2 минуты. После предварительной гипервентиляции легких эта задержка дыхания увеличивается до 3-4 минут. Однако затяжное, например, ныряние после гипервентиляции таит в себе серъезную опасность. Быстрое падение оксигенации крови может вызвать внезапную потерю сознания, а в этом состоянии пловец (даже опытный) под влиянием стимула, вызванного ростом парциального напряжения углекислоты в крови, может вдохнуть воду и захлебнуться (утонуть).
Итак, в заключение лекции я должен Вам напомнить, что здоровое дыхание это - через нос, как можно реже, с задерджкой во время вдоха и, особенно, после него. Удлиняя вдох, мы стимулируем работу симпатического отдела вегетативной нервной системы, со всеми вытекающими отсюда последствиями. Удлиняя выдох, мы удерживаем больше и дольше в крови углекислоту. А это оказыавает положительное влияние на тонус кровеносных сосудов (снижает его), со всеми вытекающими отсюда последствиями. Благодаря этому кислород может в такой ситуации пройти в самые отдаленные сосуды микроциркуляции, препятствуя нарушению их функции и развитию многочисленных заболеваний. Правильное дыхание - это профилактика и лечение большой группы заболеваний не только дыхательной системы, но и других органов и тканей! Дышите на здоровье!
Структурно-функциональная организация дыхательного центра. Регуляция вентиляции легких.
Ритмическая последовательность вдоха и выдоха, изменение характера дыхательных движений в зависимости от состояния организма (покой, работа различной интенсивности, эмоциональные проявления и т. д.) обусловлены наличием дыхательного центра, расположенного в продолговатом мозге. Дыхательный центр - совокупность нейронов, обеспечивающих деятельность аппарата дыхания и его приспособление к изменяющимся условиям внешней и внутренней среды.
Дыхательный центр - парное, симметрично расположенное образование, в состав которого входят вдыхательная и выдыхательная части.
Предполагают, что в продолговатом мозге локализованы центры вдоха, выдоха и судорожного дыхания. В верхней части моста мозга (варолиев мост) находится пнеймотаксический центр, который контролирует деятельность расположенных ниже центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III-IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудных сегментов спинного мозга (III-XII).
Дыхательный центр обладает автоматией. В нем автоматически возникает ритмическое возбуждение с частотой 14-16 раз в минуту из-за обменных процессов в самих нейронах, которые очень чувствительны к недостатку кислорода. Т.к. дыхательная мускулатура образована поперечнополосатой мышечной тканью, человек способен произвольно изменять вентиляцию легких. Это возможно, т.к. деятельность дыхательного центра продолговатого мозга находится под контролем коры больших полушарий.
В стенке легких имеются механорецепторы, которые возбуждаются при достижении легкими определенной степени растяжения при вдохе. От этих рецепторов поступают импульсы в дыхательный центр, взывая расслабление дыхательных мышц. В дыхательном центре ритмично возникает возбуждение и торможение.
Показатели и регуляция вентиляции легких в условиях измененной газовой среды (недостаток кислорода, избыток углекислого газа, изменённое атмосферное давление) и при физической нагрузке.
У тренированных людей при напряженной мышечной работе объем легочной вентиляции возрастает до 50—100 л/мин по сравнению с 5—8 л в состоянии относительного физиологического покоя. Повышение минутного объема дыхания при физической нагрузке связано с увеличением глубины и частоты дыхательных движений. У тренированных людей, в основном, изменяется глубина дыхания, у нетренированных — частота дыхательных движений.
При физической нагрузке увеличивается концентрация в крови и тканях углекислого газа и молочной кислоты, которые стимулируют нейроны дыхательного центра как гуморальным путем, так и за счет нервных импульсов, поступающих от сосудистых рефлексогенных зон. Активность нейронов дыхательного центра обеспечивается потоком нервных импульсов, поступающих от клеток коры головного мозга, обладающих высокой чувствительностью к недостатку кислорода и к избытку углекислого газа.
Одновременно возникают приспособительные реакции в сердечно-сосудистой системе. Увеличиваются частота и сила сердечных сокращений, повышается АД, расширяются сосуды работающих мышц и суживаются сосуды других областей.
ЗАНЯТИЕ № 10: «Физиология дыхания. Регуляция вентиляции легких. Функциональная система поддержания параметров газового гомеостаза»
Дыхательный центр - совокупность нейронов, расположенных в разных отделах ЦНС, обеспечивающих координированную деятельность мышц и приспособление дыхания к условиям внешней и внутренней среды.
2. Понятие об автоматии дыхательного центра, ее значение.
Автоматия дыхательного центра поддерживается и изменяется под влиянием импульсов от дыхательных мышц, сосудистых рефлексогенных зон, различных интеро- и экстерорецепторов, а также под влиянием многих гуморальных факторов (рН крови, содержание углекислого газа и кислорода в крови и др).
Способность дыхательного центра к автоматии впервые обнаружена И.М. Сеченовым (1882) в опытах на лягушках в условиях полной деафферентации животных. В этих экспериментах, несмотря на то что афферентные импульсы не поступали в ЦНС, регистрировались колебания потенциалов в дыхательном центре продолговатого мозга.
Об автоматии дыхательного центра свидетельствует опыт Гейманса с изолированной головой собаки. Ее мозг был перерезан на уровне моста и лишен различных афферентных влияний (были перерезаны языкоглоточный, язычный и тройничный нервы). В этих условиях к дыхательному центру не поступали импульсы не только от легких и дыхательных мышц (вследствие предварительного отделения головы), но и от верхних дыхательных путей (вследствие перерезки названных нервов). Тем не менее у животного сохранились ритмические движения гортани. Этот факт можно объяснить только наличием ритмической активности нейронов дыхательного центра.
7.Укажите виды хеморецепторов и их значение в регуляции вентиляции дыхания
Центральными (медуллярными) хеморецепторами, постоянно участвующими в регуляции дыхания, называют нейрональные структуры в продолговатом мозге, чувствительные к напряжению СО2 и кислотно-щелочному состоянию омывающей их межклеточной мозговой жидкости. Хемочувствительные зоны имеются на переднебоковой поверхности продолговатого мозга около выходов подъязычного и блуждающего нервов в тонком слое мозгового вещества на глубине 0.2-0.4 мм. Медуллярные хеморецепторы постоянно стимулируются ионами водорода в межклеточной жидкости ствола мозга, концентрация которых зависит от напряжения СО2 в артериальной крови. Спинномозговая жидкость отделена от крови гемато-энцефалическим барьером, относительно непроницаемым для ионов Н+ и НСО3, но свободно пропускающим молекулярный СО2. При повышении напряжения СО2 в крови он диффундирует из кровеносных сосудов головного мозга в спинномозговую жидкость, в результате чего, в ней накапливаются ионы Н+, которые стимулируют медуллярные хеморецепторы. При повышении напряжения СО2 и концентрации водородных ионов в жидкости, омывающей медуллярные хеморецепторы, увеличивается активность инспираторных и падает активность экспираторных нейронов дыхательного центра продолговатого мозга. В результате этого, дыхание становится более глубоким и вентиляция легких растет, главным образом, за счет увеличения объема каждого вдоха. Напротив, снижение напряжения СО2 и подщелачивание межклеточной жидкости ведет к полному или частичному исчезновению реакции увеличения объема дыхания на избыток СО2 (гиперкапнию) и ацидоз, а также к резкому угнетению инспираторной активности дыхательного центра вплоть до остановки дыхания.
Периферические хеморецепторы,воспринимающие газовый состав артериальной крови, расположены в двух областях:
2) Месте деления (бифуркация) общей сонной артерии (каротидный синус),
т.е. в тех же зонах, что и барорецепторы, реагирующие на изменения кровяного давления. Однако, хеморецепторы представляют собой самостоятельные образования, заключенные в особых тельцах — клубочках или гломусах, которые находятся вне сосуда. Афферентные волокна от хеморецепторов идут: от дуги аорты — в составе аортальной ветви блуждающего нерва, а от синуса сонной артерии — в каротидной ветви языкоглоточного нерва, так называемом нерве Геринга. Первичные афференты синусного и аортального нерва проходят через ипсилатеральное ядро солитарного тракта. Отсюда хеморецептивные импульсы поступают к дорсальной группе дыхательных нейронов продолговатого мозга.
Артериальные хеморецепторывызывают рефлекторное увеличение легочной вентиляции в ответ на снижение напряжения кислорода в крови (гипоксемию). Даже в обычных (нормоксических) условиях эти рецепторы находятся в состоянии постоянного возбуждения, которое исчезает только при вдыхании человеком чистого кислорода. Уменьшение напряжения кислорода в артериальной крови ниже нормального уровня вызывает усиление афферентации из аортальных и синокаротидных хеморецепторов.
Хеморецепторы каротидного синуса. Вдыхание гипоксической смеси ведет к учащению и увеличению регулярности импульсов, посылаемых хеморецепторами каротидного тельца. Повышению напряжения СО2 артериальной крови и соответствующему подъему вентиляции также сопутствует рост импульсной активности, направляемой в дыхательный центр от хеморецепторов каротидного синуса. Особенность роли, которую играют артериальные хеморецепторы в контроле за напряжением углекислоты, состоит в том, что они ответственны за начальную, быструю, фазу вентиляторной реакции на гиперкапнию. При их денервации указанная реакция наступает позднее и оказывается более вялой, так как развивается в этих условиях лишь после того, как повысится напряжение СО2 области хемочувствительных мозговых структур.
Гиперкапническая стимуляцияартериальных хеморецепторов, подобно гипоксической, носит постоянный характер. Эта стимуляция начинается при пороговом напряжении СО2 20-30 мм рт.ст и, следовательно, имеет место уже в условиях нормального напряжения СО2 в артериальной крови (около 40 мм рт.ст.).
8. Дать классификацию рецепторов механоцептивного контура регуляции вентиляции легких
Читайте также: