Ультраструктура животных и растительных клеток. Клеточные мембраны.
Добавил пользователь Владимир З. Обновлено: 28.01.2026
Довольно сложное строение мембран — важнейшая особенность эукариотических клеток.
Мембраны клетки находятся вокруг цитоплазмы, ядра, митохондрий и пластид. С их помощью образуются лабиринты эндоплазматической сети (ретикулума) и кучка сплющенных пузырьков, формирующих комплекс Гольджи. Мембраны также окружают большие вакуоли, лизосомы и пероксисомы.
Структуры, окруженные мембранами, соответствуют определенным компрантментам — они принимают участие в определенных метаболических процессах и циклах. Отдельные химические реакции происходят на самих мембранах. Речь идет о световой реакции фотосинтеза в хлоропластах и окислительном фосфорилировании в процессе дыхания в митохондриях. Строение мембраны клетки невозможно представить без рецепторных участков, которые отвечают за распознавание внешних стимулов, в частности, гормонов и прочих химических веществ, поступающих из вне или других частей организма.
Сохранение различий между содержимым клетки и внешней средой — заслуга плазматической мембраны, которая окружает каждую клетку. Поэтому существование клетки без мембраны невозможно.
Плазматическая мембрана (плазмалема) — наиболее постоянная, основная и универсальная для всех типов клеток система поверхностного аппарата.
По своему строению плазматическая мембрана — тонкая (от 6 до 10 нм), но плотная пленка, покрывающая все клетку, образованная упорядоченно расположенными молекулами белков и фосфолипидов. Нековалентные связи удерживают их вместе.
Плазматическая мембрана состоит из двух рядов липидов, молекулы которых располагаются таким образом, что неполярные гидрофобные концы находятся в глубине мембраны, а полярные гидрофильные концы — направлены на внешнюю и внутреннюю среду.
Липидный слой не является сплошным: отдельные места мембраны пронизаны белковыми молекулами, которые образуют гидрофильные поры. Через эти поры проходят водорастворимые вещества. Другие молекулы белка размещаются на внешней и внутренней сторонах мембраны.
На поверхности всех эукариотических клеток есть углеводы, которые, в основном, ковалентно связаны с мембранными белками (гликопротеидами), но иногда — с липидами (гликолипидами). Масса углеводов в плазматической мембране — от 2 до 10%.
Плазматические мембраны клетки — это подвижные структуры. Молекулы белков, липидов, полисахаридов, ионы кальция, калия, натрия и другие вещества — основная составляющая мембраны клетки. Эти вещества способны быстро перемещаться в плоскости мембраны и менять в ней свое расположение. Миграция всех этих веществ происходит при помощи диффузии или путем поглощения кислорода.
Также вне зависимости от строения плазматические мембраны отличаются динамичностью — в случае повреждения они способны к быстрому восстановлению. Кроме того, они могут растягиваться и сжиматься при активном движении клеток.
В различных видах клеток мембраны различаются по:
- химическому составу;
- содержанию в них белков, липидов, гликопротеидов;
- характеру рецепторов, которые содержатся в мембранах.
Отсюда и индивидуальность типов клеток, определяемая гликопротеидами. Факторы внешней среды распознаются разветвленными цепями гликопротеидов, выступающими над клеточной мембраной, через реакции клеток на их действие.
Яйцеклетка и сперматозоид, образующие одно целое, могут узнавать друг друга по гликопротеидам на поверхности клетки. Взаимное узнавание является важным этапом, предшествующим процесс оплодотворение. Это явление наблюдается в ходе дифференциации тканей. При помощи распознающих участков на плазмалеме клетки, похожие по строению, могут правильно ориентироваться относительно друг друга и образовывать, в результате, ткани.
Узнавание тесно связано с регулированием транспорта молекул и ионов через мембрану и иммунологическим ответом, в котором гликопротеиды играют роль антигенов.
Так функционирование сахаров осуществляется в виде информационных молекул — похоже, как у белков и нуклеиновых кислот.
В строении мембран выделяют специфические рецепторы, транспортирующие электроны, ферментные белки. Благодаря белкам обеспечивается транспорт некоторых молекул в клетку и обратно, осуществляется структурная связь цитоскелета с клеточными мембранами. Белки выполняют функции рецепторов — они получают или преобразуют химические сигналы окружающей среды.
Еще одна жизненно важная функция плазматической мембраны, обусловленная ее четкой структурной организацией и упорядоченностью — полупроницаемость или способность выборочно пропускать в клетку и выпускать из нее разнообразные молекулы и ионы. Все это формирует в клетке определенную концентрацию ионов и обеспечивает процесс осмоса.
Мембраны, похожие по строению и являющиеся основными структурными элементами клетки, ограничивают почти все ее органеллы. Это не просто физические мембраны, а динамически функциональная поверхность. На мембранах органелл происходят различные биохимические процессы:
- поглощение неорганических и органических веществ;
- синтез АТФ;
- превращение энергии квантов света в ходе фотосинтеза и др.
Транспорт веществ через плазматическую мембрану
Основная функция плазматической мембраны — регулирование обмена различными веществами, который происходит между клеткой и окружающей средой.
Есть 4 варианта поступления веществ в клетку и выхода из нее:
- Диффузия.
- Активный транспорт.
- Эндо- и экзоцитоз.
- Осмос.
В растворах перемещение растворенных веществ происходит в следующем направлении: из участка, где отмечается высокая концентрация, в участок, где отмечается низкая концентрация. Поток веществ в сторону с меньшей концентрацией (транспорт по градиенту концентраций) сохраняется до момента выравнивания концентрации в обоих участках.
Диффузия
Диффузия или диффузное перемещение веществ — это перемещение веществ, на которое оказывает влияние градиент концентраций.
Размер молекулы и ее относительная растворимость в жирах определяют скорость диффузии через мембрану. Чем молекула меньше, тем она быстрее растворяется в липидах и легче диффундирует через мембрану.
Без проблем растворяются в липидном слое мембраны и проходят через нее, оказываясь с другой стороны, небольшие неполярные молекулы. Маленькие незаряженные молекулы, такие как этанол, углекислый газ, мочевина, быстро проходят через мембрану. А вот глюкоза, кислоты, глицерины и аминокислоты проходят через мембрану медленно.
Несмотря на то, что молекулы воды плохо растворяются в жирах, вода легко проходит через липидный слой. Все потому, молекулы воды небольших размеров.
Липидные слои плазматической мембраны являются серьезным препятствием для попадания в клетку для всех заряженных молекул (ионов). И их размер не имеет значения.
Специфические белки отвечают за перенос различных полярных молекул (ионы, сахара, нуклеотиды, аминокислоты и прочие метаболиты) через клеточные мембраны. Эти белки получили название мембранных транспортных белков.
Определенный белок отвечает за транспорт каждого конкретного химического соединения. Специализированные транспортные белки могут соединяться с молекулой или ионом, не затрачивая на это энергию, то есть, пассивно, и транспортировать их через мембрану по градиенту концентрации.
Такой процесс получил название облегченной диффузии. Он является основным механизмом выборочной проницательности мембран.
Активный транспорт
В отличие от облегченной диффузии, активный транспорт веществ происходит против градиентов их концентрации. Осуществляется переход веществ из участка низкой концентрации в участок высокой концентрации. В этом случае без затрат энергии не обойтись (ее источником выступает АТФ): она используется для перенесения протонов или неорганических ионов через мембрану.
Все это можно продемонстрировать на примере. Расход энергии обеспечивает попадание катиона калия в клетки корня растения в условиях, когда его концентрация в почвенном растворе в 100 раз меньше, чем в клеточном соке. А вот катион натрия, который растению не особо нужен, может выводиться в окружающую среду даже в том случае, если его концентрация в почве, в которой находятся корни, достаточно высока.
Активное поглощение свойственно только ионам питательных элементов. Из этого следует, что клетка отличается определенной выборочной способностью к разным ионам. Прочие ионы попадают в клетку согласно градиенту их электрохимического потенциала и проницательности мембраны.
Эндоцитоз и экзоцитоз
Макромолекулы попадают внутрь клетки при помощи эндоцитоза. Речь идет о белках, полисахаридах, полинуклеотидах.
Есть 2 типа эндоцитоза:
- Фагоцитоз, основанный на поглощении твердых частичек. Клетки окружают капли жидкости с плотными частичками и втягивают их в цитоплазму. Там под действием ферментов происходит их расщепление до фрагментов, которые клетка может усвоить.
- Пиноцитоз, основанный на поглощении жидкостей. Процесс поглощения жидкостей похож на фагоцитоз. В месте, где капля и клетка касаются, образуется впячивание в виде канальца, который заполняется жидкостью. Далее происходит его отшнуровывание и попадание в цитоплазму — там мембранные стенки пузырька распадаются, и содержимое освобождается. В процессе пиноцитоза клетки могут поглощать и большие молекулы, и ионы, у которых нет возможности проникнуть через мембрану в силу маленьких размеров пор.
Фагоцитоз и пиноцитоз обеспечивают питание гетеротрофных протист, защитные реакции высших организмов (лейкоциты поглощают чужеродные частички) и транспорт веществ (в клетках почечных канальцев происходит всасывание белков из первичной мочи).
Экзоцитоз — процесс, противоположный эндоцитозу. В ходе него происходит вывод содержимого пузырька из клетки во внешнюю среду.
К примеру, клетки, продуцирующие гормон инсулин, запаковывают его в пузырьки внутри клетки. Потом пузырьки сливаются с плазматической мембраной и открываются наружу — происходит освобождение инсулина.
Осмос
Осмос — это диффузия воды через полупроницаемую мембрану, которая происходит за счет разницы или градиента концентраций внутри клетки и во внешней среде.
Путем осмоса клетка обычно поглощает воду.
При помещении клетки в гипотонический раствор, можно будет наблюдать градиент водного потенциала. Концентрации воды снаружи клетки превысит концентрацию внутри клетки. По этой причине поступление воды внутрь клетки происходит по градиенту ее собственной концентрации — мембрана выборочно пропускает только молекулы воды.
При помещении клетки в гипертонический раствор, который более концентрированный снаружи, вода будет выходить из клетки под действием осмотических сил.
В гипертоническом растворе происходит смарщивание эритроцитов, уменьшение вакуоли в мембране растительной клетки, отставание цитоплазмы от клеточной стенки (плазмолиз). Таким образом растения вянут.
Если будет действовать определенное внешнее давление (осмотическое давление), то движение воды через мембрану при наличии градиента концентрации можно остановить. Это объясняется желанием молекул воды пройти через полупроницаемую мембрану и уровнять концентрацию с обеих сторон мембраны.
Силы, потраченные на препятствование проникновению воды в раствор (наружу или внутрь) через полупроницаемую мембрану, зависят от концентрации раствора (чем она выше, тем больше нужно сил).
По этой причине осмотическое давление раствора с большей концентрацией выше, чем разбавленного. В первом случае раствор сильнее поглощает воду из окружающего раствора.
Определить осмотическое давление можно путем определения числа частичек в одной единице объема растворителя.
Уровень концентрации ионов и молекул разнообразных соединений в растительной клетке будет выше, чем в окружающей ее естественной среде, в частности, в почве. Поэтому клетка развивает всасывающую силу, которая позволяет ей всасывать воду снаружи.
Происходит набухание клетки и формирование внутреннего гидростатического давления — оно направлено на клеточную стенку. Его называют тургорным давлением. Противоположность тургорного давления — механическое давление клеточной стенки (оболочки), направленное внутрь клетки, которое имеет такую же величину, что и тургорное.
Таким образом, мы рассмотрели строение и функции плазматической мембраны.
Клетка
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
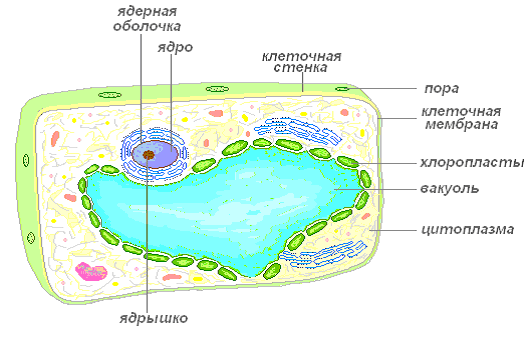
Строение растительной клетки
Строение и функции органоидов растительной клетки
Клеточная стенка или плазматическая мембрана
Бесцветная, прозрачная и очень прочная
Пропускает в клетку и выпускает из клетки вещества.
Клеточная мембрана полупроницаемая
Густое тягучее вещество
В ней располагаются все другие части клетки
Находится в постоянном движении
Ядро (важная часть клетки)
Округлое или овальное
Обеспечивает передачу наследственных свойств дочерним клеткам при делении
Центральная часть клетки
Сферической или неправильной формы
Принимает участие в синтезе белка
Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок
Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке.
По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль
Используют световую энергию солнца и создают органические из неорганических
Форма дисков, отграниченных от цитоплазмы двойной мембраной
Образуются в результате накопления каротиноидов
Жёлтые, оранжевые или бурые
Содержатся в корнях, клубнях, луковицах
Состоит из двух мембран (наружная и внутренняя) с порами
Отграничивает ядро от цитоплазмы
Даёт возможность осуществляться обмену между ядром и цитоплазмой
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.
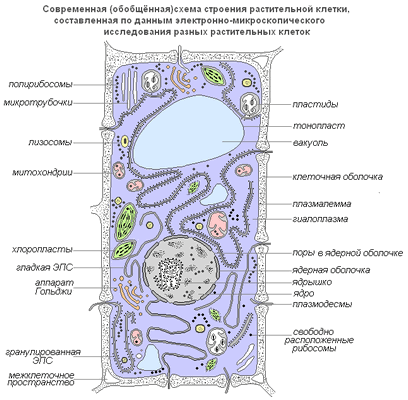
Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования - органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.
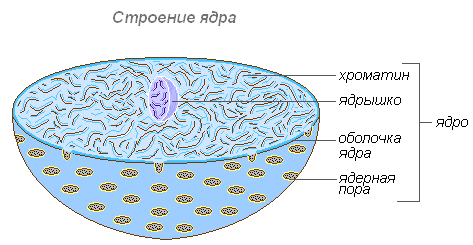
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
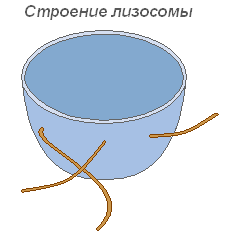
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
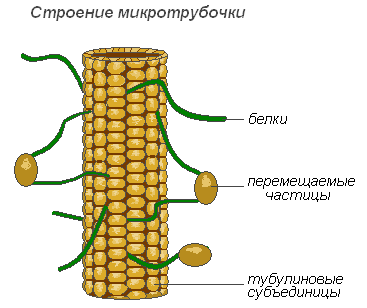
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.
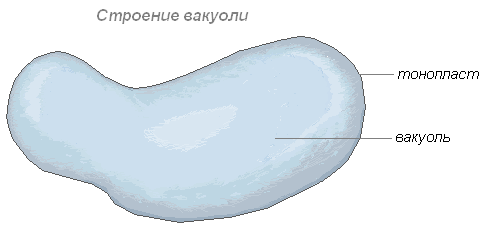
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.
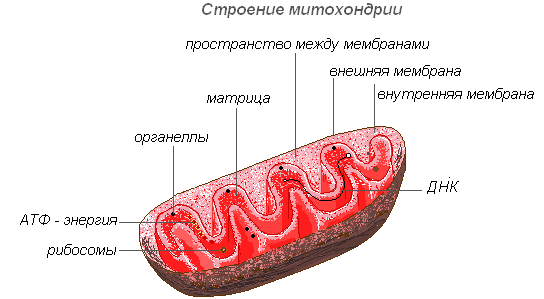
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.
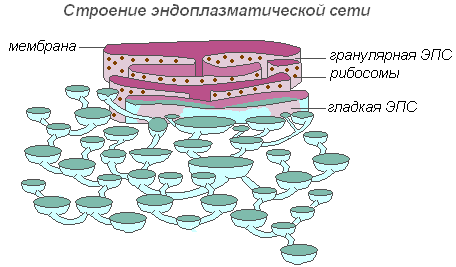
Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.
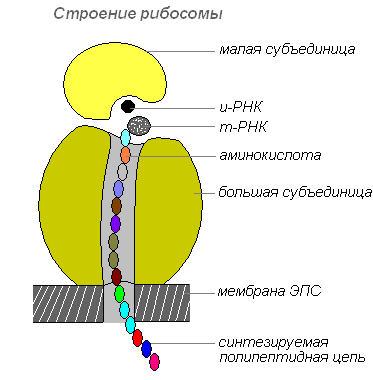
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
БИОЛОГИЯ Том 1 - руководство по общей биологии - 2004
Тонкая структура клетки, выявляемая при помощи электронного микроскопа, называется ультраструктурой. На рис. 5.10 и 5.11 представлена ультраструктура обобщенных клеток, животной и растительной, а на рис. 5.12 и 5.13 электронные микрофотографии типичной животной и типичной растительной клеток. Приведена и характеристика всех показанных здесь компонентов клеток.
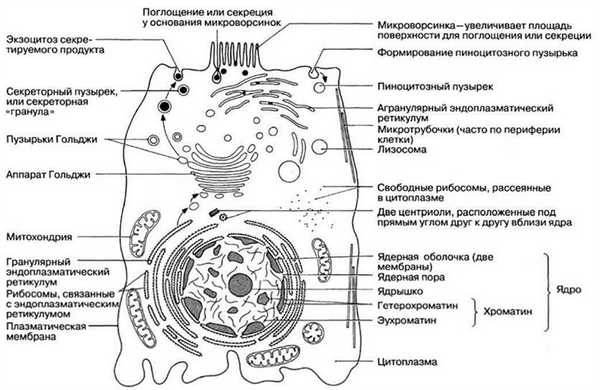
Рис. 5.10. Ультраструктура обобщенной животной клетки, выявляемая при помощи электронного микроскопа. Для простоты показана лишь часть гранулярного эндоплазматического ретикулума с присоединенными к нему рибосомами и некоторое количество свободных рибосом.
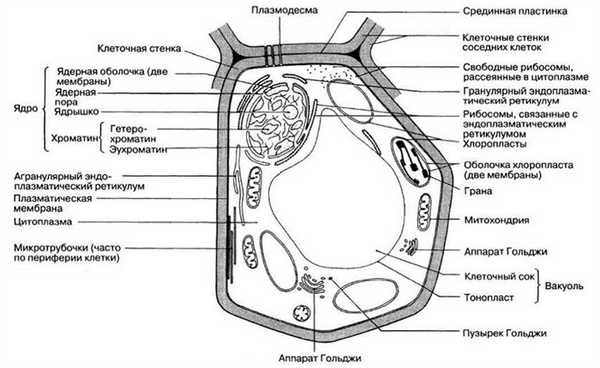
Рис. 5.11. Ультраструктура обобщенной растительной клетки, выявляемая при помощи электронного микроскопа.
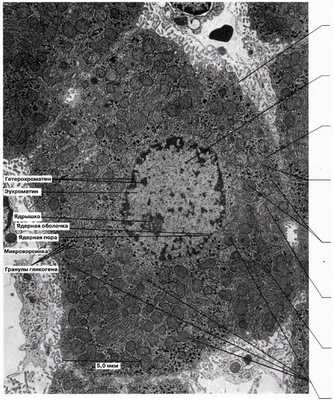

Рис. 5.12. Электронная микрофотография тонкого среза типичной животной клетки из печени крысы — гепатоцита (х9600).
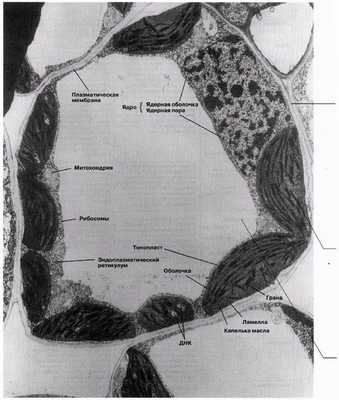
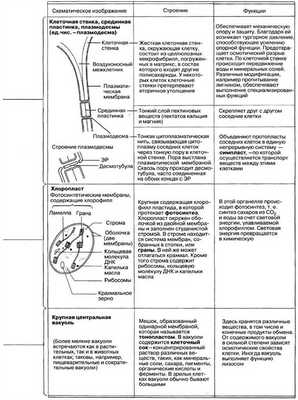
Рис. 5.13. Электронная микрофотография тонкого среза типичной растительной клетки — клетки мезофилла листа (х15 000).
5.1. Сопоставьте рис. 5.1 и 5.2 с рис. 5.10 и 5.11. Какие новые структуры выявляет электронный микроскоп в сравнении со световым микроскопом?
5.2. Сравнив рис. 5.1 и 5.2 с рис. 5.10 и 1.11, отметьте, какие структуры а) имеются в растительных клетках и отсутствуют в животных и б) имеются в животных клетках и отсутствуют в растительных?
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Строение и функции биологических мембран
Биологические мембраны - общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки, гетерогенные молекулы (гликопротеины, гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем - рецепторов, ферментов, транспортных механизмов - помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
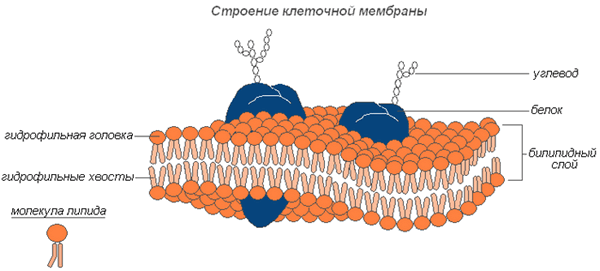
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими, непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными, погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
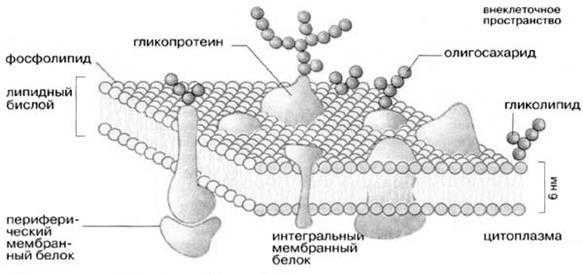
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).
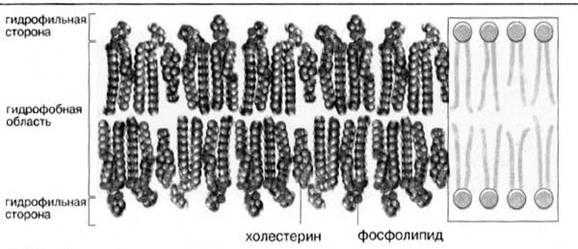
Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами. Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.
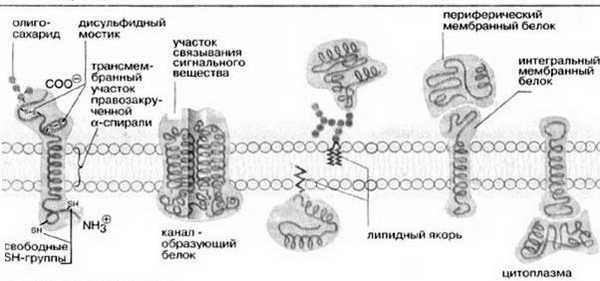
Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний - фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа + из клетки в окружающую среду, а ионы К + - внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса.
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная - с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое - фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин, состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны, окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них - фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов. Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система - ламеллы. Ламеллы образуют или уплощенные пузырьки - тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
Ультраструктура прокариотической клетки
Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов —прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер — пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток — бактериальные споры;
клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет — до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.
Рис. 1. Схема строения прокариотической клетки:
/ — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — мезосомы; 5 — цитоплазма; 6 — нуклеоид; 7 — плазмида;
8 — рибосомы и полисомы; 9 — жгутики; 10 — пили; 11 — гранулы гликогена; 12 — липидные капельки; 13 — гранулы волютина; 14 — включения серы
Поверхностные структуры. Капсула — это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) — это прямые цилиндрические образования белковой природы длиной 0,3—10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики — органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3—12 мкм и толщиной 10—30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и располо-
жение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2—30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка — один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм и L-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01—14 мкм. Она представляет собой плотную эластичную структу -
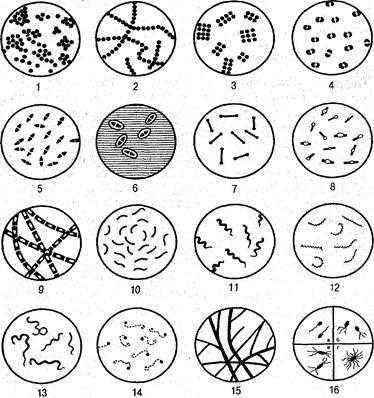
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ — стафилококки; 2 — стрептококки; 3 — сарцины; 4 — гонококки;
5— пневмококки; 6— капсулы пневмококков; 7— коринебактерии дифтерии; 8 — клостридии; 9 — бациллы; 10 — вибрионы; 11 — спириллы; 12 — трепонсмы; — боррелии; 14 — лептоспиры; 15— актиномицеты; 16 — расположение жгутиков: а — монотрихи; б — лофотрихи; в — амфитрихи; г — перитрихи
ру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер — пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы. Цитоплазматическая мембрана (ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7—13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15—30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50—70%). В мембране имеются также углеводы (2—5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы — впячивания ЦМ — смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазма заполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70—80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характер-
ными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид — ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2—3 10 в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды — необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы — органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S (так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц — 30S и 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80S рибосом.
Споры (эндоспоры) бактерий — покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключен-
Иной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора — это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий — бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения. Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других — запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы — гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.
Читайте также:
- Расстройство пищевого поведения: причины, симптомы и лечение
- Алкогольная анозогнозия. Формы анозогнозии у больных алкоголизмом
- Двурогая матка. Причины и диагностика
- Появление систолического шума при инфаркте миокарда. Разрыв межжелудочковой перегородки, папиллярных мышц
- Общие сведения о заболевании перикарда