Строение гликокаликса - капсулы прокариот
Добавил пользователь Владимир З. Обновлено: 29.01.2026
• Надмембранные комплексы клеток состоят из структур, расположенных над плазматической мембраной. В частности, это клеточная стенка клеток растений, грибов и прокариот, а также гликокаликс животных клеток.
• Строение клеточной стенки. Вы уже знаете, что в клетках бактерий, грибов и растений плазматическая мембрана снаружи покрыта более или менее плотной клеточной стенкой. У растений она включает собранные в пучки нерастворимые в воде волокна полисахарида целлюлозы (рис. 16.1) и своеобразный каркас. В состав клеточной стенки растений входят и другие полисахариды, например пектин, гемицеллюлоза и др.
В зависимости от типа тканей и выполняемых ими функций в состав клеточной стенки растений могут входить и другие вещества: липиды, белки, неорганические соединения (SiO2, карбонаты и фосфаты кальция и т. п.). Например, оболочки клеток пробки или сосудов с возрастом просачиваются жирообразным веществом суберином. Вследствие этого содержимое клетки отмирает, что способствует выполнению опорной или проводящей функций. Клеточные стенки способны одревесневать, когда промежутки между волокнами целлюлозы заполняет полисахарид лигнин, повышающий прочность стенок.
Рис. 16.1. Клеточная стенка растительной клетки
Рис. 16.2. 1. Явление плазмолиза: при высокой концентрации солей во внеклеточной среде вода выходит из клетки, внутриклеточное давление уменьшается и цитоплазма отслаивается от клеточной стенки. 2. Явление деплазмолиза: при низкой концентрации солей во внеклеточной среде вода поступает в клетку и внутриклеточное давление возрастает
У разных групп грибов основу клеточной стенки также составляют полисахариды. Кроме целлюлозы, это может быть азотсодержащий полисахарид хитин (вспомните животных, в состав внешнего скелета которых входит это вещество), гликоген и т. п. В состав клеточных стенок некоторых грибов могут входить темные пигменты (меланины) и другие соединения. Строение клеточных стенок прокариот вы изучите позже.
• Функции клеточной стенки. Основная функция клеточной стенки - опорная - поддержание формы клетки. Другая функция - защитная - защита внутреннего содержимого клетки от механических повреждений. Еще одна функция - транспортная - перемещение воды и других соединений в клетку и из нее.
Проницаемость клеточных стенок растений проявляется в явлениях плазмолиза и деплазмолиза (рис. 16.2). Например, если растительную клетку поместить в раствор с концентрацией солей более высокой, чем в цитоплазме, то вода будет выходить из клетки наружу. Это вызывает явление плазмолиза (от греч. плазма - лепка, образование и лизис - растворение) - отслоение пристеночного слоя цитоплазмы от клеточной стенки (рис. 16.2, 1). Если же клетку внести в раствор солей с более низкой концентрацией, чем в цитоплазме, то наблюдают обратный процесс: вода будет поступать в клетку, вследствие чего возрастет внутриклеточное давление. Этот процесс называют деплазмолизом (от лат. де - префикс, обозначающий отличие) (рис. 16.2, 2). Наблюдать явления плазмолиза и деплазмолиза учащиеся, изучающие биологию на академическом уровне, смогут самостоятельно, выполнив лабораторную работу № 4 (см. лабораторный практикум на с. 109).
Клеточная стенка имеет много мелких отверстий - пор, соединенных с подобными образованиями соседних клеток. Через них содержимое соседних клеток сообщается с помощью цитоплазматичних тяжей - плазмодесм (см. рис. 15.8, 3). Как вы помните, плазмодесмы расположены в окруженных плазматической мембраной канальцах, проходящих через оболочки клеток. Диаметр пор составляет 30-60 нм. По оси канальца проходит цилиндрическая трубка меньшего диаметра, соединенная с эндоплазматической сетью обеих клеток. Считают, что плазмодесмы служат для транспорта веществ непосредственно из клетки в клетку.
• Гликокаликс. Клетки животных не имеют твердой клеточной стенки; над их плазматической мембраной расположен гликокаликс (от лат. гликис - сладкий и каллюм - толстая кожа) (рис. 16.3). Этот поверхностный слой толщиной в несколько десятков нанометров состоит из соединений белков и липидов с углеводами.
Гликокаликс обеспечивает непосредственную связь клеток с окружающей средой. В его состав входят рецепторные молекулы, способные воспринимать раздражители окружающей среды. Он также участвует в избирательном транспорте веществ (пропускает или не пропускает) молекулы, в зависимости от их размеров, заряда и т. п. Благодаря наличию ферментов гликокаликс может участвовать в примембранном пищеварении - расщеплении соединений, которые расположены снаружи вблизи поверхности клетки. В итоге клетка потребляет эти продукты расщепления. Кроме того, гликокаликс обеспечивает межклеточные связи у многоклеточных животных.
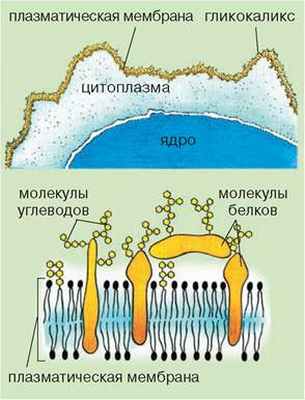
Рис. 16.3. Схема строения гликокаликса
• Подмембранные комплексы клеток. К ним относятся разнообразные структуры белковой природы: микронити (микрофиламенты) и микротрубочки, которые образуют цитоскелет (рис. 16.4, 1). Цитоскелет выполняет опорную функцию, а также соединяет все компоненты клетки: ее поверхностный аппарат, структуры цитоплазмы, ядро. Элементы цитоскелета способствуют закреплению в определенном положении и перемещению органелл в клетке.
Микронити, или микрофиламенты, - тонкие нитевидные структуры, диаметром 4-7 нм, которые состоят из сократительных белков, преимущественно актина (рис. 16.4, 3). Они пронизывают цитоплазму и могут образовывать сплетение под плазматической мембраной. Пучки микронитей прикреплены одним концом к определенной структуре (например, плазматической мембране), а вторым - к другой (органелле и т.п.). Микрофиламенты участвуют в изменении формы клетки, например при амебоидном движении, процессах поступления в клетку и выводе из нее различных соединений. При делении некоторых клеток животных они образуют особое сократительное кольцо, разделяющее цитоплазмы дочерних клеток.
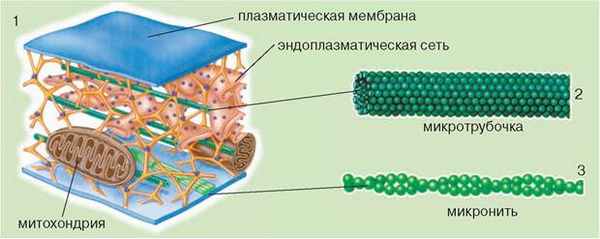
Рис. 16.4. Схема строения цитоскелета (1), микротрубочек (2) и микронитей (3)
Микротрубочки - полые цилиндрические структуры диаметром 10-25 нм, образованные преимущественно белком тубулином (рис. 16.4, 2). Они участвуют в формировании веретена деления эукариотических клеток, входят в состав ресничек, жгутиков и т. п. Микротрубочки обеспечивают перемещение органелл и макромолекул в клетке. При этом пучки микротрубочек одним концом прикрепляются к одной структуре или молекуле, а вторым - к другой.
У некоторых одноклеточных животных структура цитоскелета усложнена. Например, у радиолярий сферическая капсула из органического вещества разделяет цитоплазму на внутреннюю и внешнюю части. Во внешней части содержатся жировые включения. Они уменьшают плотность содержимого клетки и облегчают «парение» этих организмов в толще воды. Во внутренней части цитоплазмы содержатся ядро и много других органелл. Кроме того, в цитоплазме находятся внутриклеточные структуры из неорганических веществ - SiO2 или SrSO4. Они могут иметь вид дырчатых сфер, заключенных одна в другую, корон, радиально расположенных игл и т. п. (рис. 16.5). К ним прикреплены микронити, другим концом связанные с плазматической мембраной и способные к сокращению. Эти микронити регулируют плотность животных; во время шторма радиолярии опускаются вглубь, а в спокойную погоду питаются другими одноклеточными, в частности водорослями, вблизи поверхности.
В клетках многих одноклеточных животных (инфузорий, эвглен и т. п.) к подмембранным комплексам относится пелликула (от лат. пеллис - кожа). Она состоит из структур, расположенных в уплотненном внешнем слое цитоплазмы. Так, у инфузорий в состав пелликулы входят уплощенные цистерны из органического вещества, которые вместе образуют мозаичную структуру. Внутри таких цистерн могут находиться дополнительные опорные комплексы из белков или пропитанных карбонатом кальция полисахаридных пластинок. Пелликула придает прочность оболочке клетки, обеспечивая относительное постоянство ее формы.
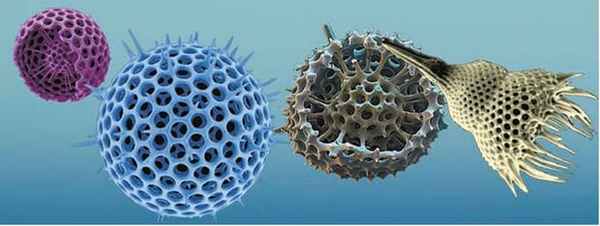
Рис. 16.5. Внутриклеточные скелеты радиолярий
Важными функциями подмембранного комплекса клеток являются обеспечение цитоза, движения (образование псевдоподий) и т. п. При этом микротрубочки и микронити удлиняются или укорачиваются.
Ключевые термины и понятия. Плазмолиз, деплазмолиз, гликокаликс, микрофиламенты, микротрубочки, пелликула.
Кратко о главном
- Над плазматической мембраной расположен поверхностный аппарат. В клетках грибов и растений плазматическую мембрану снаружи окружает клеточная стенка. Клеточная стенка защищает внутреннее содержимое клетки и поддерживает ее форму. Через нее осуществляется транспорт воды и других соединений.
- Надмембранные комплексы клеток животных представлены гликокаликсом. Он обеспечивает связь клетки с окружающей средой и между клетками у многоклеточных животных.
- К подмембранным комплексам относится цитоскелет, образованный из белковых структур - микронитей (микрофиламентов) и микротрубочек. Элементы цитоскелета способствуют закреплению органелл в определенном положении или их перемещению.
Вопросы для самоконтроля
1. Что такое клеточная стенка? Каковы ее функции? 2. Какие особенности строения клеточной стенки у растений и грибов? 3. Что такое плазмолиз и деплазмолиз? 4. Что такое гликокаликс? Каковы его функции? 5. Что такое цитоскелет? Каковы его функции? 6. Что такое пелликула? Какое ее значение?
Подумайте
1. Что общего и отличного в строении и функциях клеточной стенки и гликокаликса? 2. Одноклеточная водоросль хламидомонада и представитель одноклеточных животных эвглена зеленая имеют много общих особенностей строения клетки и процессов жизнедеятельности. Какие именно признаки позволяют отнести хламидомонаду к царству Растения, а эвглену зеленую - к царству Животные?
Строение гликокаликса - капсулы прокариот
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Обозначения:
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
Один из основных признаков прокариотической клетки— отсутствие внутреннего разделения, обеспечиваемого элементарными мембранами. По сути, единственная мембранная система прокариотической клетки — ЦПМ, входящая в состав клеточной оболочки, часто сложно устроенная и проникающая поперечными складками глубоко в цитоплазму. В составе любой стабильной бактериальной клетки выделяют условные компартменты — поверхностные структуры, клеточную оболочку и цитоплазму.
Поверхностные структуры бактерий
Основные поверхностные структуры бактериальной клетки— капсула, жгутики и микроворсинки. Их наличие — относительно стабильный признак, используемый для идентификации бактерий.
Капсула бактерий
Клеточную оболочку многих бактерий окружает слой аморфного, сильно обводнённого вещества. Этот покров выполняет важные функции: делает оболочку клетки (состоящей из клеточной стенки и ЦПМ) более плотной и прочной, предохраняет бактерии от воздействия бактерицидных факторов, обеспечивает адгезию на различных субстратах, может содержать запасы питательных веществ.
Организация капсул бактерий. Основную роль в организации капсул бактерий играет ЦПМ. Выделяютмикрокапсулы(выявляемые только при электронной микроскопии в виде слоя мукополисахаридных микрофибрилл) имакрокапсулы(обнаруживают при световой микроскопии). У некоторых бактерий полимеры клеточной оболочки, выделяемые наружу, свободно располагаются вокруг неё, образуя слизистый слой. Капсулированные бактерии могут превращаться в бескапсульные варианты и, поскольку первые образуют мукоидные или гладкие (S), а бескапсульные — шероховатые (R) колонии, это явление известно как S- и R-диссоциация. Капсула и слизистый слой не препятствуют поступлению и выходу различных веществ из бактериальной клетки, а также плохо удерживают красители.
Окраска капсул бактерий. Для облегчения микроскопирования капсулы можно сделать видимыми, проведя негативнуюокраску мазка по Бурри-ГинсуилиХиссулибо с помощью реакциинабухания по Нойфельду(см. ниже).
Состав капсул бактерий. В состав большинства бактериальных капсул входят сложные полисахариды. Капсулы также могут содержать соединения азота (например, у пневмококков капсула состоит из полисахаридов, глюкозамина и глюкуроновой кислоты), но могут и не содержать азот (например, капсулы лейконостоков состоят из декстрана, левулана, фруктозана и других полимеризованных моносахаров). Капсулы некоторых болезнетворных бактерий (например, Bacillus anthracis) формируют полисахариды и полипептиды, образованные мономерами D-глутаминовой кислоты. Поскольку D-аминокислоты устойчивы к воздействию протеаз, такая капсула лучше защищает бактерию от ферментативных воздействий фагоцитов.
Антигенные свойства капсул бактерий. Капсульные Аг (К-Аг) многих патогенных бактерий проявляют выраженные иммуногенные свойства (например, вакцины против пневмококковых и менингококковых инфекций готовят из материала капсулы). Связывание AT с капсулой изменяет её светопреломляющие характеристики, вызывая кажущееся набухание капсулы, видимое при соответствующих условиях освещения под микроскопом и известное какреакция Нойфельда.
Строение гликокаликса - капсулы прокариот
• У многих прокариот наружная поверхность содержит слой, обогащенный полисахаридами, который называется капсула или слизистый слой
• Предполагается, что капсула или слизистый слой предотвращает высыхание бактерий, обеспечивает их связывание с рецепторами клеток хозяина при попадании в организм, а также помогает бактериям избежать атаки со стороны его иммунной системы
• Формирование капсулы у Е. coli происходит по крайней мере по одному из четырех различных путей
• У многих прокариот, наряду с капсулой или вместо нее, существует S-слой. представляющий собой внешнее белковое покрытие обладающее кристаллическими свойствами
Большинство прокариот, если не все, поверх клеточной стенки образуют гликокаликс или капсулу. Термин гликокаликс приложим к клеткам как прокариот, так и эукариот; и используется для обозначения смеси внеклеточных полисахаридов и белков. В случае прокариотических клеток, между различными типами гликокаликса существуют довольно произвольные различия. Полисахариды, ковалентно связанные с клеткой, называются капсулой.
Непрочно связанные полисахариды, способные отщепляться от клетки, называются «слизистым слоем» или внеклеточными полисахаридами. Состав капсулы значительно варьирует, однако обычно в состав ее входят полисахариды, включая полиспирты и аминосахара, протеогликаны и гликопротеины. Толщина и степень пластичности этого слоя варьируют в зависимости от его химического состава. Полисахариды капсулы образуются при объединении моносахаридов в длинные цепи.
Поскольку любой моносахарид способен соединиться с другим моносахаридом, существует большое разнообразие капсулярных полисахаридов. Серологические свойства капсулярных ингредиентов можно использовать для выяснения отличий ее состава у родственных организмов. Например, у Е. coli описано более 80 различных капсулярных полисахаридов или К-антигенов. Известно, что штаммы, экспрессирующие некоторые К-антигены, связаны с определенными инфекциями.
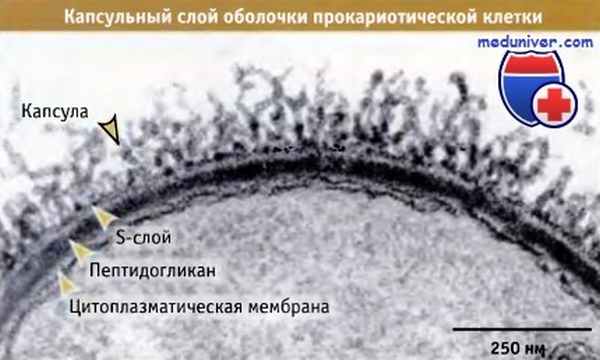
У большинства прокариот капсула представляет собой наружный слой клеточной оболочки.
На этой микрофотографии представлена оболочка Bacillus anthracis.
Капсульный слой выполняет несколько функций, включая защиту клеток от высыхания, фагоцитов, детергентов и бактериофагов. Защита от высыхания играет важную роль в переносе инкапсулированных микроорганизмов от одного хозяина к другому. Капсула выполняет важную роль, обеспечивая адгезию клеток к тканям хозяина и к внешним поверхностям. Капсулы также обеспечивают связывание с другими прокариотами, что приводит к образованию биопленки. Например, Pseudomonas aeruginosa продуцирует огромные количества альгинатов, которые способствуют образованию биопленки в легких.
Такая прочная пленка из альгината также способствует защите патогенных микробов от антибиотиков и защитных систем организма хозяина.
Капсула играет важную роль не только при инфицировании организма хозяина патогенным микроорганизмом Р. aeruginosa. Например, основным вирулентным фактором Bacillus anthracis, вызывающего сибирскую язву, оказалась капсула. Капсула В. anthracis состоит из полимера D-глутаминовой кислоты. Это единственная капсула прокариот, состоящая из пептидов. Материал капсулы продуцируется in vivo, и штаммы В. anthracis, неспособные синтезировать капсулу, характеризуются ослабленной патогенностью. Считается, что, поскольку материал капсулы В. anthracis обладает слабой иммуногенностью, она защищает патогенный микроорганизм от атаки иммунной системы организма хозяина. Капсула также оказывает ингибирующее действие на связывание белков комплемента, тем самым позволяя микробу существовать в системе кровообращения хозяина.
Капсула также является основным фактором вирулентности Mycobacterium tuberculosis. У Mycobacterium капсула обеспечивает связывание с макрофагами, что служит критическим шагом в проникновении бактерии в организм хозяина. Поскольку макрофаги являются фагоцитами, которые захватывают и разрушают микробы, последние выработали различные механизмы, позволяющие им выживать находясь внутри макрофагов. Для Mycobacterium капсула играет критическую роль в одном из таких механизмов.
Во-первых, она обеспечивает связывание с CR3 рецептором макрофагов, что вызывает сигнальный каскад, приводящий к «безопасному» пути инфекции. Также она играет важную роль в нейтрализации активных кислород-радикалов, присутствующих в макрофагах и способных разрушить микроб. Это помогает объяснить, каким образом энкапсулированные Mycobacteria избегают атаки иммунной ситемы и персистируют в организме хозяина, в то время как бескапсульные штаммы обладают меньшей вирулентностью. В действительности, бескапсульные штаммы Mycobacterium используются для изготовления вакцины, обеспечивающей иммунитет к тубекулезу.
Биогенез капсулы изучен на нескольких видах бактерий, однако, вероятно, лучше всего на E. coli. По генетическим и биохимическим критериям капсулы этой бактерии подразделяются на четыре группы. Особенно подробно исследован биогенез групп 1 и 2. Транспорт капсул этих групп к поверхности клетки, вероятно, происходит в местах тесного контакта между цитоплазматической и наружной мембранами. В транслокации капсул, относящихся к первой группе, участвует высокомолекулярный комплекс, который образует Wza, представляющий собой липопротеид наружной мембраны. Предполагается, что Wza, представляющий собой белок, формирующий цилиндр (b-barrel), вместе с Wzc образует канал, через который происходит секреция предшественников капсулы. Эта система секреции функционально и генетически близка к Usher белку наружной мембраны, РарС.
При участии белков KpsE и KpsD, компоненты капсул группы 2 мигрируют через наружную мембрану в обоих направлениях. Белок KpsE связывается с плазматической мембраной через свой N-концевой домен, в то время как большая часть белка находится в периплазматическом пространстве. Хотя KpsE не пронизывает наружную мембрану, С-концевой участок его молекулы связан с ней. Функционирование такого секреторного аппарата зависит от взаимной близости цитоплазматической и наружной мембран. Хотя функция белка KpsD до конца не выяснена, известно, что он представляет собой периплазматический белок, который, возможно, необходим для мобилизации белка KpsE на наружной мембране. Биогенез капсул, относящихся к группам 3 и 4, изучен недостаточно.
У многих бактерий и археев на наружной поверхности находится белковая структура, которая называется поверхностный слой (S-слой). Ее можно рассматривать как альтернативу капсуле или как дополнение к ней. S-слои образуются из одного белка или фрагментов гликопротеинов, которые в процессе самосборки формируют кристаллическую решетку. Структура белка не является консервативной, однако часто он содержит много остатков кислых и гидрофобных аминокислот. Иногда, главным образом у археев и грамположительных бактерий, обнаруживаются гликозилированные белки. Особенно характерно наличие множественных сайтов гликозилирования.
Гликозильные цепи состоят из 20-50 идентичных субъединиц, содержащих набор сахарных остатков. На рисунке ниже представлен препарат S-слоя, приготовленный с использованием метода «замораживания-травления» (видна поверхность, обладающая гексагональной симметрией, и поперечный срез через S-слой). Обычно толщина S-слоя составляет от 5 до 25 нм. Наружная поверхность его довольно гладкая, а внутренняя выглядит сморщенной в тех местах, где слой взаимодействует с более глубоко лежащими слоями. Сборка S-слоя находится под контролем клеточной оболочки, которая взаимодействует с такими молекулами, расположенными на поверхности, как теихоевые кислоты (у грамположительных бактерий) или липополисахариды (у грамотрицательных бактерий). Каким образом сборка и позиционирование S-слоя связаны с капсулой (в случае когда существуют два слоя), пока не ясно.
Если у археев пептидогликан или эквивалентный компонент отсутствует, то S-слой представляет собой основной компонент клеточной оболочки. Хотя белки S-слоя составляют значительную часть тотального клеточного белка, их функции пока не вполне выяснены. Многие штаммы прокариот, культивируемые в лаборатории, утратили способность формировать S-слой, однако прекрасно растут. Возможно, существует некий смысл не формировать этот слой в условиях, когда он не требуется.

У некоторых прокариот снаружи вместо капсульного слоя находится S-слой.
Слева: электронная микрофотография, иллюстрирующая гексагональную симметрию наружной поверхности S-слоя.
Справа: электронная микрофотография поперечного среза клеточной оболочки, содержащей S-слой.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Строение бактериальной клетки
В составе любой стабильной бактериальной клетки выделяют условные компартменты — поверхностные структуры (капсула, жгутики, микроворсинки), клеточную стенку и цитоплазму. Бактериальная клетка представляет собой сложную живую высокоупорядоченную систему с определенным обменом веществ. Она включает целый ряд морфологически и химически обособленных структур, каждая из которых выполняет определенную функцию.
Капсула. Многие бактерии покрыты слоем аморфного, сильно обводненного вещества — капсулой, состоящей на 98% из воды. Это можно увидеть с помощью светового микроскопа (рис. 3.9), если добавить к препарату красители, которые в капсулу не проникают (нигрозин, Конго красный, китайская тушь). Основную роль в организации капсул бактерий играет ЦПМ. Выделяют макрокапсулы (до 10 мкм, обнаруживают при световой микроскопии), превышающие величину самой бактериальной клетки, микрокапсулы (меньше 0,2 мкм, выявляют только при электронной микроскопии в виде слоя мукополисахаридных микрофибрилл), слизистый слой (у некоторых бактерий полимеры клеточной стенки выделяются наружу) и растворимую слизь (многие компоненты капсулы выделяются в окружающую среду в виде слизи, особенно обильное образование слизи наблюдается у многих микроорганизмов в тех случаях, когда среда содержит сахарозу).
Капсулированные бактерии могут превращаться в бескапсуль- ные варианты, первые образуют мукоидные, или гладкие (5), а бескапсульные — шероховатые (К) колонии. Это явление известно как явление S- и ^-диссоциации. Капсула и слизистый слой не препятствуют поступлению и выходу различных веществ из бактериальной клетки, а также плохо удерживают красители.
Химический состав капсулы у разных бактерий неодинаков. В состав большинства бактериальных капсул входят сложные высокомолекулярные полисахариды, нередко содержащие еще ацетильные и аминные группы и полипептиды, образованные а-глю-
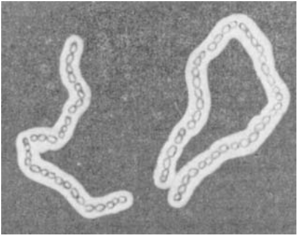
Рис. 3.9. Капсулы бактерий
Цепочки клеток Bacillus megaterium суспендированы в растворе туши. На темном фоне капсулы выделяются как светлые зоны. Фазовый контраст (увел. 1000 х)
таминовой кислотой. Капсульные полисахариды помимо глюкозы содержат аминосахара, рамнозу, уроновые кислоты, пирови- ноградную и уксусную кислоты. Например, капсулы Leuconostoc mesenteroides (представитель гетероферментативных молочнокислых бактерий, который быстро превращает тростниковый сахар в декстрановый студень) состоят из декстрана, левулана, фруктоза- на и других полимеризованных моносахаров. Капсулы же Bacillus anthracis (возбудитель сибирской язвы) содержат полисахариды и полипептиды, образованные мономерами .D-глютаминовой кислоты. Поскольку D-аминокислоты устойчивы к воздействию протеаз, такая капсула лучше защищает бактерию от ферментативных воздействий фагоцитов. Капсулы туберкулезных бактерий состоят из липидов и воскоподобных веществ (Mycobacterium tuberculosis).
Образование капсулы зависит от условий развития и питания бактерий. У некоторых видов капсула образуется при большом количестве углеводов и низком содержании белков в питательной среде. Некоторые патогены образуют капсулу при размножении в организме животных или человека и не образуют ее на обычных питательных средах (возбудитель сибирской язвы). Некоторые исследователи полагают, что капсула представляет собой набухшую наружную часть клеточной стенки, а другие считают ее продуктом выделения, отличающимся по своей химической природе от наружной клеточной стенки. Вторая точка зрения очевидна, так как иногда достаточно тщательно промыть препарат, для того чтобы освободить бактериальные клетки от капсулы.
Функции капсулы. У бактерий она является защитным приспособлением против неблагоприятного воздействия внешней среды (пересыхание), фагов, токсических веществ и др. Капсульные антигены (К — Аг) многих патогенных бактерий проявляют выраженные иммуногенные свойства. Обладание капсулой придает некоторым патогенным бактериям резистентность к фагоцитозу и тем самым повышает их вирулентность для подопытных животных. В природных условиях некоторые представители нормальной микрофлоры человека вырабатывают так называемый глико- каликс, состоящий из длинных полисахаридных фибрилл и обеспечивающий клеткам тесное прилипание (Steptococcus mutans — вызывающий кариес (разрушение) зубов). Эти полисахариды откладываются на поверхности зубов и служат матриксом, в котором накапливаются кислые продукты брожения, главным образом молочная кислота.
Влагалища (или чехлы). Вокруг тела некоторых нитчатых бактерий образуются вследствие затвердения наружных слоев оболочки (явление, обратное капсулообразованию) особые чехлы, или влагалища, или кожистые пленки. Например, Acetobacter aceti subsp. xylinum выделяет целлюлозу, которая объединяет клетки в прочную кожистую пленку («Mycoderma aceti»). Клетки Sarcina ventriculi склеиваются целлюлозой в агрегаты правильной формы. Целлюлоза в этих случаях служит связующим веществом и по своей структуре и функции отличается от веществ капсулы. У железобактерий в чехлах отлагается гидроокись железа, вследствие чего они приобретают прочность. На дне озер и болот образуются отложения руды, состоящей из остатков железобактерий.
Зооглей. Обычно капсула окружает от одной до трех-четырех микробных клеток, иногда же слизистая масса образуется вокруг большого количества микробных клеток. Такое скопление микробов, заключенных в общую капсулу, называется зооглеем.
Читайте также: