Регуляция секреторной функции ( секреции ) тонкой кишки. Местные рефлексы.
Добавил пользователь Алексей Ф. Обновлено: 28.01.2026
Регуляция деятельности желез тонкой кишки осуществляется местными нервно-рефлекторными механизмами, а также гуморальными влияниями и ингредиентами химуса. Механическое раздражение слизистой оболочки тонкой кишки вызывает выделение жидкого секрета с малым содержанием ферментов. Местное раздражение слизистой кишки продуктами переваривания белков, жиров, соляной кислотой, панкреатическим соком вызывает отделение кишечного сока, богатого ферментами. Усиливают кишечное сокоотделение ГИП, ВИП, мотилин. Гормоны энтерокринин и дуокринин, выделяемые слизистой оболочкой тонкой кишки, стимулируют соответственно секрецию либеркюновых и бруннеровых желез. Тормозное действие оказывает соматостатин.
27. Диагностика кишечной непроходимости
27. Диагностика кишечной непроходимости При осмотре живота необходимо обратить внимание на его конфигурацию, общий или местный метеоризм и асимметрию передней брюшной стенки.В начальном периоде кишечной непроходимости в первую очередь растягивается петля кишки,
40. Лечение врожденной кишечной непроходимости
40. Лечение врожденной кишечной непроходимости Предоперационную подготовку проводят строго индивидуально. У новорожденных с высокой кишечной непроходимостью длительность и качество предоперационной подготовки зависят от тяжести состояния, времени поступления в
46. Клиника острой кишечной инвагинации
46. Клиника острой кишечной инвагинации Клиническая картина острой инвагинации зависит от уровня внедрения кишки, возраста ребенка и сроков, прошедших с начала заболевания.Клиническая картина слепо-ободочной и подвздошно-ободочной инвагинации. Заболевание начинается
47. Диагностика острой кишечной инвагинации
47. Диагностика острой кишечной инвагинации Бесконтрастная обзорная рентгенография брюшной полости не оказывает значительной помощи в установлении диагноза инвагинации и позволяет выявить только некоторые косвенные признаки непроходимости (отсутствие газа в толстой
48. Лечение острой кишечной инвагинации
48. Лечение острой кишечной инвагинации Основной принцип лечения инвагинации кишок - возможно ранняя дезинвагинация. Хирургическая тактика и методы лечебных мероприятий зависят от локализации и формы внедрения, сроков поступления и возраста ребенка. Существует два
51. Лечение ранней спаечной кишечной непроходимости
51. Лечение ранней спаечной кишечной непроходимости Лечение ранней спаечной непроходимости требует индивидуального подхода в зависимости от общего состояния ребенка, развития основного заболевания, сочетанных осложнений и сроков, прошедших от момента первой
55. Клиника и диагностика паралитической кишечной непроходимости
55. Клиника и диагностика паралитической кишечной непроходимости I стадия возникает непосредственно вслед за оперативным вмешательством. В этой стадии пареза отсутствуют органические изменения в интрамуральных сплетениях; микроциркуляторные изменения в кишечной
56. Лечение паралитической кишечной непроходимости
56. Лечение паралитической кишечной непроходимости Лечение паралитической непроходимости кишечника слагается из коррекции системных нарушений гомеостаза и борьбы с локальными проявлениями пареза.Мероприятия по лечению локальных проявлений пареза условно можно
Модификация рациона при кишечной диспепсии
Модификация рациона при кишечной диспепсии Если у больного выявляется та или иная диспепсия , то в зависимости от ее вида в рацион питания больного вводят необходимые коррективы.Бродильная диспепсияБродильная диспепсия (dyspepsia fermentativa) обычно обусловлена преобладанием в
Регуляция функций желез внутренней секреции
Регуляция функций желез внутренней секреции Регуляция деятельности желез внутренней секреции осуществляется нервными и гуморальными факторами. Нейроэндокринные зоны гипоталамуса, эпифиз, мозговое вещество надпочечников и другие участки хромаффинной ткани
Регуляция желудочной секреции
Регуляция желудочной секреции Железы желудка вне процесса пищеварения выделяют только слизь и пилорический сок. Отделение желудочного сока начинается при виде, запахе пищи, поступлении ее в ротовую полость. Процесс желудочного сокоотделения можно разделить на
Регуляция секреции поджелудочной железы
Регуляция секреции поджелудочной железы Регуляция поджелудочной экзокринной секреции осуществляется нервными и гуморальными механизмами. Блуждающий нерв усиливает секрецию поджелудочной железы. Симпатические нервы уменьшают количество секрета, но усиливают синтез
Открытие кишечной микрофлоры
Открытие кишечной микрофлоры Бактериальная природа инфекционных болезней была открыта Луи Пастером лишь в конце XIX в. Незадолго до этого Пастер открыл и бактериальную природу гниения белковых продуктов и возможность стерилизации продуктов питания нагреванием.
Восстановление кишечной микрофлоры
Восстановление кишечной микрофлоры Кишечную микрофлору можно восстановить при помощи чесночной простокваши. Это молочнокислый продукт, приготовленный с использованием чеснока. Такая простокваша прекрасно восстанавливает микрофлору кишечника, устраняя последствия
73. Состав и свойства кишечного сока. Регуляция секреции кишечного сока.
Сок в тонком кишечнике представляет собой бесцветную, мутноватую жидкость. Имеет щелочную реакцию. За сутки у человека выделяется около 2-х литров кишечного сока. Из органических веществ в нем содержатся кристаллы холестерина, аминокислоты, мочевина и другие вещества; из неорганических - соли Na, К и небольшое количество углекислых солей. В кишечном соке содержатся ферменты, действующие на БЖУ.
К ферментам, действующим на белки, относятся:
Эрепсин - это сложный (комплексный) фермент, состоящий из разных протеолитических ферментов, он расщепляет альбумины и белки до аминокислот.
Нуклеаза - действует на сложные белки, расщепляя их до нуклеиновых кислот.
К ферментам, действующим на жиры, относятся:
Липаза - расщепляет жиры, которые достигли кишечника еще не расщепленными.
К ферментам, действующим на углеводы, относятся:
Образование ферментов кишечного сока отличается по своему типу от ферментов других пищеварительных желёз. Секреторные клетки слюнных желёз, желёз желудка, поджелудочной железы, выделяя секрет, сохраняют свою целостность. Этот тип называется морфостатическим. Отделение ферментов кишечного сока сопровождается гибелью железистой клетки, эти клетки отторгаются и выделяются вместе с калом во внешнюю среду - морфоникротический тип секреции.
Выделение кишечного сока происходит непостоянно, а в связи с действием на слизистую кишечника поступившего химуса. Этот химус раздражает клетки механически и своими физико-химическими свойствами.
Регуляция секреции кишечного сока.
Основную роль в регуляции сокоотделения слизистой оболочки кишечника играют местные рефлексы, осуществляемые энтеральной НС. Механическое раздражение усиливает секрецию жидкой части кишечного сока и не изменяет содержания в нем ферментов. Стимуляция хеморецепторов продуктами переваривания белков и жиров вызывает секрецию сока, богатого ферментами. При раздражении блуждающего нерва увеличивается содержание ферментов в кишечном соке. Такой же эффект оказывают АЦХ и холиномиметики. Раздражение чревного нерва угнетает отделение сока.
Во время приема пищи наблюдается рефлекторное усиление секреции бруннеровых желез ДПК и рефлекторное торможение остального железистого аппарата, что предотвращает избыточную выработку сока и его ферментов (сверх того количества секрета, которое обусловлено местным раздражение химуса рецепторов кишки).
Гастроинтестинальные гормоны, вырабатываемые эндокринными элементами слизистой оболочки кишки под влиянием химуса (дуокринин, энтерокинин, ГИП, ВИП и мотилин), а также гормоны коры надпочечников (кортизон, дезоксикортикостерон) стимулируют кишечную секрецию, подкрепляя функцию местных рефлекторных механизмов, и только один гормон - соматостатин - оказывает тормозное влияние на кишечное сокоотделение.
Секреторная функция тонкой кишки
Ежедневно в тонком кишечнике образуется до 2 л секрета (кишечныйсок) с pH от 7,5 до 8,0. Источники секрета — железы подслизистой оболочки двенадцатиперстной кишки (бруннеровы железы) и часть эпителиальных клеток ворсинок и крипт.
Бруннеровыжелезысекретируют слизь и бикарбонаты. Слизь, выделяемая бруннеровыми железами, защищает стенку двенадцатиперстной кишки от действия желудочного сока и нейтрализует соляную кислоту, поступающую из желудка.
Эпителиальныеклеткиворсинокикрипт(рис. 22-8). Их бокаловидные клетки секретируют слизь, а энтероциты выделяют в просвет кишки воду, электролиты и ферменты.
Ферменты. На поверхности энтероцитов в ворсинках тонкой кишки находятсяпептидазы(расщепляют пептиды до аминокислот),дисахаридазысукраза, мальтаза, изомальтаза и лактаза (расщепляют дисахариды на моносахариды) икишечнаялипаза(расщепляет нейтральные жиры до глицерина и жирных кислот).
Регуляциясекреции. Секрециюстимулируютмеханическое и химическое раздражение слизистой оболочки (местные рефлексы), возбуждение блуждающего нерва, гастроинтестинальные гормоны (особенно холецистокинин и секретин). Секрецию тормозят влияния со стороны симпатической нервной системы.
Секреторная функция толстой кишки. Крипты толстой кишки выделяют слизь и бикарбонаты. Величину секреции регулируют механическое и химическое раздражение слизистой оболочки и локальные рефлексы энтеральной нервной системы. Возбуждение парасимпатических волокон тазовых нервов вызывает увеличение отделения слизи с одновременной активацией перистальтики толстой кишки. Сильные эмоциональные факторы могут стимулировать акты дефекации с периодическим выделением слизи без фекального содержимого («медвежья болезнь»).
Переваривание пищи
Белки, жиры и углеводы в пищеварительном тракте превращаются в продукты, способные всасываться (пищеварение, переваривание). Продукты пищеварения, витамины, минералы и вода проходят сквозь эпителий слизистой оболочки и поступают в лимфу и кровь (всасывание). Основу пищеварения составляет химический процесс гидролиза, осуществляемый пищеварительными ферментами.
Углеводы. В пище содержатсядисахариды(сахароза и мальтоза) иполисахариды(крахмалы, гликоген), а также другие органические соединения углеводного характера.Целлюлозав пищеварительном тракте не переваривается, так как у человека нет ферментов, способных её гидролизовать.
Ротоваяполостьижелудок.-Амилаза расщепляет крахмал до дисахарида — мальтозы. За короткое время пребывания пищи в ротовой полости переваривается не более 5% всех углеводов. В желудке углеводы продолжают перевариваться в течение часа, прежде чем пища полностью перемешается с желудочным соком. За этот период до 30% крахмалов гидролизуется до мальтозы.
Тонкаякишка.-Амилаза панкреатического сока заканчивает расщепление крахмалов до мальтозы и других дисахаридов. Содержащиеся в щёточной каёмке энтероцитов лактаза, сахараза, мальтаза и-декстриназа гидролизуют дисахариды. Мальтоза расщепляется до глюкозы; лактоза — до галактозы и глюкозы; сахароза — до фруктозы и глюкозы. Образовавшиеся моносахариды всасываются в кровь.
Желудок. Пепсин, активный при pH от 2,0 до 3,0, превращает 10-20% белков в пептоны и некоторое количество полипептидов.
Тонкаякишка(рис. 22-8)
Ферменты поджелудочной железы трипсин и химотрипсин впросветекишкирасщепляют полипептиды на ди- и трипептиды, карбоксипептидаза отщепляет аминокислоты от карбоксильного конца полипептидов. Эластаза переваривает эластин. В целом образуется немного свободных аминокислот.
На поверхности микроворсинок каёмчатых энтероцитов в двенадцатиперстной и тощей кишке находится трёхмерная густая сеть — гликокаликс, в котором расположены многочисленные пептидазы. Именно здесь эти ферменты осуществляют так называемое пристеночноепищеварение. Аминополипептидазы и дипептидазы расщепляют полипептиды на ди- и трипептиды, а ди- и трипептиды превращают в аминокислоты. Затем аминокислоты, дипептиды и трипептиды легко транспортируются внутрь энтероцитов через мембрану микроворсинок.
В каёмчатых энтероцитах имеется множество пептидаз, специфичных для связей между конкретными аминокислотами; в течение нескольких минут все оставшиеся ди- и трипептиды превращают в отдельные аминокислоты. В норме более 99% продуктов переваривания белков всасывается в виде отдельных аминокислот. Очень редко всасываются пептиды.
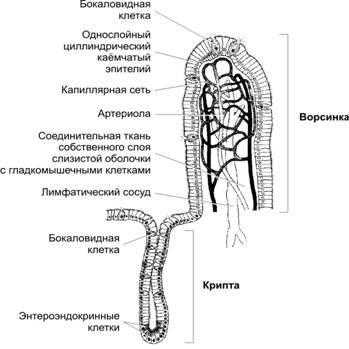
Рис.22-8.Ворсинкаикриптатонкогокишечника[11]. Слизистая оболочка покрыта однослойным цилиндрическим эпителием. Каёмчатые клетки (энтероциты) участвуют в пристеночном пищеварении и всасывании. Панкреатические протеазы в просвете тонкого кишечника расщепляют поступающие из желудка полипептиды на короткие пептидные фрагменты и аминокислоты с последующим их транспортом внутрь энтероцитов. Расщепление коротких пептидных фрагментов до аминокислот происходит в энтероцитах. Энтероциты передают аминокислоты в собственный слой слизистой оболочки, откуда аминокислоты поступают в кровеносные капилляры. Связанные с гликокаликсом щеточной каёмки дисахаридазы расщепляют сахара до моносахаридов (главным образом, глюкозы, галактозы и фруктозы), которые всасываются энтероцитами с последующим выходом в собственный слой и поступлением в кровеносные капилляры. Продукты пищеварения (кроме триглицеридов) после всасывания через капиллярную сеть в слизистой оболочке направляются в воротную вену и далее в печень. Триглицериды в просвете пищеварительной трубки эмульгируются жёлчью и расщепляются панкреатическим ферментом липазой. Образовавшиеся свободные жирные кислоты и глицерин поглощают энтероциты, в гладкой эндоплазматической сети которых происходит ресинтез триглицеридов, а в комплексеГольджи— формирование хиломикронов — комплекса триглицеридов и белков. Хиломикроны подвергаются экзоцитозу на боковой поверхности клетки, проходят через базальную мембрану и поступают в лимфатические капилляры. В результате сокращения ГМК, расположенных в соединительной ткани ворсинки, лимфа продвигается в лимфатическое сплетение подслизистой оболочки. Кроме энтероцитов, в каёмчатом эпителии присутствуют бокаловидные клетки, вырабатывающие слизь. Их количество нарастает от двенадцатиперстной к подвздошной кишке. В криптах, особенно в области их дна, расположены энтероэндокринные клетки, вырабатывающие гастрин, холецистокинин, желудочный ингибирующий пептид, мотилин и другие гормоны.
Жирынаходятся в пище преимущественно в виде нейтральных жиров (триглицеридов), а также фосфолипидов, холестерола и эфиров холестерола. Нейтральные жиры входят в состав пищи животного происхождения, их значительно меньше в растительной пище.
Желудок. Липазы расщепляют менее 10% триглицеридов.
Тонкаякишка
Переваривание жиров в тонкой кишке начинается с превращения крупных жировых частиц (глобул) в мельчайшие глобулы — эмульгированиежиров(рис. 22-9А). Этот процесс начинается в желудке под влиянием перемешивания жиров с желудочным содержимым. В двенадцатиперстной кишке жёлчные кислоты и фосфолипид лецитин эмульгируют жиры до размеров частиц в 1 мкм, увеличивая общую поверхность жиров в 1000 раз.
Панкреатическая липаза расщепляет триглицериды на свободные жирные кислоты и 2-моноглицериды и способна в течение 1 минуты переварить все триглицериды химуса, если они находятся в эмульгированном состоянии. Роль кишечной липазы в переваривании жиров невелика. Накопление моноглицеридов и жирных кислот в местах переваривания жиров останавливает процесс гидролиза, но этого не происходит, потому что мицеллы, состоящие из нескольких десятков молекул жёлчных кислот, удаляют моноглицериды и жирные кислоты в момент их образования (рис. 22-9А). Мицеллы холатов транспортируют моноглицериды и жирные кислоты к микроворсинкам энтероцитов, где они всасываются.
Фосфолипиды содержат жирные кислоты. Эфиры холестерола и фосфолипиды расщепляются специальными липазами поджелудочного сока: холестерол-эстераза гидролизует эфиры холестерола, а фосфолипаза A2расщепляет фосфолипиды.
Кислотозависимые состояния у детей /Под ред. В.А.Таболина. - Регуляция желудочной секреции
Различают базальную желудочную секрецию, которая имеет место между приемами пищи, и стимулированную - при приеме пищи.
Базальная секреция, вероятнее всего, обусловлена стимуляцией секреторных клеток желудка импульсами блуждающего нерва. О тонусе последнего можно судить по базальной концентрации панкреатического полипептида и так как концентрация панкреатического полипептида в сыворотке крови изменяется синхронно с изменениями уровня базальной секреции [7] можно сделать вывод, что базальная секреция контролируется главным образом тонусом блуждающего нерва.
Принято выделять три фазы желудочной секреции, наблюдаемой в ходе нормального пищеварительного процесса: мозговую (кефалическую), желудочную и кишечную.
Мозговая фаза начинается с выработки желудочного сока под действием условных рефлексов. Ожидание пищи или ее вид сопровождается выделением слюны и желудочного сока. Когда пища попадает в рот, возбуждение вкусовых и обонятельных рецепторов приводит к дополнительному, безусловно-рефлекторному, усилению секреции. Центры секреторных рефлексов лежат в промежуточном мозге, лимбической коре и гипоталамусе. Отсюда возбуждение поступает к желудку по волокнам блуждающего нерва. Ацетилхолин (АХ) действует как непосредственно на париетальные клетки [1, 6], так и опосредованно, стимулируя продукцию гастрина G-клетками. Выброс гастрина в кровь начинает стимулировать гистаминоциты, расположенные вокруг париетальных клеток, к выделению ими гистамина, который в свою очередь связывается с H2-рецепторами париетальных и главных клеток. Итогом является секреция HCl и желудочных ферментов.
Своего максимума этот процесс достигает во вторую фазу секреции - желудочную, когда гастрин выделяется в значительно больших количествах под влиянием непереваренных пептидов, находящихся в просвете желудка.
Продуцирующие гастрин G-клетки располагаются в слизистой оболочке пилорического отдела желудка, но их нет ни в области дна, ни в теле. Они являются клетками открытого типа, т.е. имеют рецепторы, открытые в просвет ЖКТ. Со стороны просвета желудка они стимулируются непереваренными пептидными молекулами и высоким рН. Стимуляторами секреции гастрина также являются аминокислоты, соединения кальция, которыми богаты продукты животного происхождения, в значительно меньшей степени стимулируют выделение гастрина жиры и углеводы. Оптимальным рН для гастриновой секреции является величина от 5 до 7, значения ниже 5 ингибируют ее, а при 1,7 она полностью подавляется. Налицо типичный контур саморегуляции по принципу отрицательной обратной связи: закисление желудочного содержимого приводит к торможению секреции гастрина и, как следствие, кислотной продукции.
Гастрин действует на париетальные клетки непосредственно и через стимуляцию Ecl-клеток, продуцирующих гистамин [2, 9].
Указанным стимулирующим воздействиям противостоит ряд тормозящих влияний. Важнейшим звеном регуляции является секреция соматостатина, который тормозит функцию париетальных, гастрин- и гистамин-продуцирующих клеток. D-клетки, продуцирующие этот пептид, располагаются как в теле желудка, так и в пилорическом отделе, однако в теле желудка они являются закрытыми (т.е. не несут на своей поверхности открытых в просвет рецепторов), а в пилорическом отделе - открытыми. Следовательно, в первом случае они реагируют лишь на эндогенные факторы, а во втором - также и на влияния со стороны просвета желудка. Данные о влиянии АХ на секрецию соматостатина (СС) противоречивы: в одних работах показано его стимулирующее влияние [5], в других - тормозящее [3]. Последнее более вероятно.
Антральные D-клетки реагируют на рН: максимальная секреция СС происходит при рН=1, а подавляется при значениях выше 3. Сам же СС тормозит секрецию гастрина [10]. Так формируется регуляторный контур гастрин-кислота-соматостатин-гастрин. Более того, на D-клетках обнаружены также стимулирующие рецепторы к гастрину [8], что обеспечивает прямую регуляцию гастрин-соматостатин. В этот же контур вносят свой вклад гастрин-релизинг-пептид, который способствует секреции СС [4], а также гастрин-ингибирующий пептид, который потенциирует выброс СС в ответ на закисление среды и таким путем подавляет секрецию гастрина и кислоты.
Выделение гастрина продолжается и в начале кишечной фазы, когда в кишку поступает свежая порция пищи, не успевшая полностью пропитаться кислотой, но уже в конце желудочной фазы секреции, когда pH в антральном отделе достигает значений ниже 3, начинаются процессы торможения желудочной секреции. В кишечную фазу при попадании в двенадцатиперстную кишку желудочного содержимого с pH ниже 4, из клеток слизистой оболочки кишки выделяется секретин, который не только стимулирует щелочную секрецию поджелудочной железы, но также подавляет выделение гастрина и желудочную секрецию.
Регуляция кишечной секреции.
Местные механизмы регуляции. Механическое раздражение слизистой оболочки тонкой кишки увеличивает выделение жидкой части сока. Химическими стимуляторами секреции тонкой кишки являются продукты переваривания белка, жира, панкреатический сок, соляная и другие кислоты. Продукты переваривания питательных веществ при местном их воздействии вызывают отделение кишечного сока, богатого ферментами.
Гуморальная регуляция. Кишечную секрецию стимулируют гастроингибирующий пептид, вазоинтестинальный пептид, тормозит ее соматостатин.
Моторика тонкой кишки.
1.маятникообразные сокращения (последовательное сокращение кольцевых и продольных мышц) способствуют перемешиванию химуса с пищеварительными соками.
2. ритмическая сегментация (кольцевые м-цы) растирание и перемешивание химуса.
3. перистальтические сокращения (одновременно продольные и кольцевые м-цы) продвижение химуса.
4. тонические суживают просвет на небольших участках.
Регуляция моторики.
Интрамуральная нервная система. На нее оказывают влияние симпатическая (тормозит сокращение) и парасимпатическая (усиливает) н.с. при сокращении предшествующего отдела последующий расслабляется благодаря реципрокной активности нейронов интрамуральной н.с.
Гуморальная регуляция. Усиливают моторику: вазопрессин, серотонин, гистамин, гастрин, холецистокинин, кислоты..
Всасывание в тонкой кишке.
Всасывание — это процесс перехода веществ с поверхности клеточных мембран и из полостей органов в кровь и лимфу. Уже примерно через 7 часов после начала процесса пищеварения продукты гидролиза питательных веществ почти полностью исчезают из полости тонкого кишечника, что связано с особенностями строения слизистой кишечника и огромными размерами всасывательной поверхности.
Процесс всасывания может осуществляться пассивно — за счет осмоса и диффузии и активно — за счет специальной насосной функции ворсинчатого аппарата.
Пристеночное пищеварение и его значение.
Вещества из полости тонкой кишки поступают в слой кишечной слизи, обладающей более высокой ферментативной активностью, чем жидкое содержимое полости тонкой кишки.
В слизистых наложениях адсорбированы ферменты из полости тонкой кишки (панкреатические и кишечные), из разрушенных энтероцитов и транспортированные в кишку из кровотока. Проходящие через слизистые наложения питательные вещества частично гидролизуются этими ферментами и поступают в слой гликокаликса, где продолжается гидролиз питательных веществ по мере их транспорта в глубь пристеночного слоя. Продукты гидролиза поступают на апикальные мембраны энтероцитов, в которые встроены кишечные ферменты, осуществляющие собственно мембранное пищеварение, в основном гидролиз димеров до стадии мономеров. Следовательно, пристеночное пищеварение последовательно идет в трех зонах: слизистых наложениях, гликокаликсе и на апикальных мембранах энтероцитов с огромным числом микроворсинок на них. Образовавшиеся в результате пищеварения мономеры всасываются в кровь и лимфу.
Пищеварение в толстой кишке.
1. сгущение содержимого вследствие всасывания воды
2. брожение за счет микрофлоры
Значение микрофлоры толстой кишки для пищеварения и функций организма.
Представлена бифидо-, лактобактериями и др.
1. конечное разложение непериваренных остатков пищи
2. инактивация и расщепление ферментов
3. подавляет патогенные микроорганизмы, предотвращает инфицирование
4. синтез вит. К и гр. В
5. обмен белков, фосфолипидов, желчных и жирных к-т, билирубина, холестерина.
Моторика толстой кишки.
Моторика толстой кишки обеспечивает резервуарную функцию — накопление содержимого, всасывание из него ряда веществ, в основном воды, продвижение его, формирование каловых масс и их удаление (дефекация).
Заполнение и опорожнение.У здорового человека контрастная масса через 3—3,5 ч после ее приема начинает поступать в толстую кишку. Она заполняется в течение 24 ч и полностью опорожняется за 48—72 ч.
Типы моторики.Содержимое слепой кишки совершает небольшие и длительные перемещения то в одну, то в другую сторону за счет медленных сокращений кишки. Сокращения толстой кишки бывают нескольких типов: малые и большие маятникообразные, перистальтические и антиперистальтические, пропульсивные. Первые четыре типа сокращений перемешивают содержимое кишки и повышают давление в ее полости, что способствует сгущению содержимого путем всасывания воды. Сильные пропульсивные сокращения возникают 3—4 раза в сутки и продвигают кишечное содержимое в направлении толстой кишки.
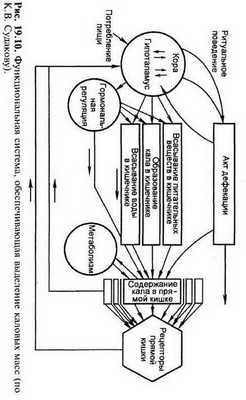
Акт дефекации.
Каловые массы удаляются с помощью акта дефекации, представляющего сложнорефлекторный процесс опорожнения дистального отдела толстой кишки через задний проход. При наполнении ампулы прямой кишки калом и повышении в ней давления до 40 - 50 см вод.ст. происходит раздражение механо- и барорецепторов. Возникшие при этом импульсы по афферентным волокнам тазового (парасимпатического) и срамного (соматического) нервов направляются в центр дефекации, который расположен в поясничной и крестцовой частях спинного мозга (непроизвольный центр дефекации). Из спинного мозга по эфферентным волокнам тазового нерва импульсы идут к внутреннему сфинктеру, вызывая его расслабление, и одновременно усиливают моторику прямой кишки.
Читайте также:
- Взаимодействие энтеральной нервной системы и иммунной системы
- Случай успешного взаимодействия стоматолога-хирурга и стоматолога-терапевта при лечении радикулярной кисты
- Околомозговое скопление жидкости
- Флегмона после операции в гинекологии
- Иммуносупрессия после трансплантации поджелудочной железы. Принципы