Строение пили, жгутиков бактерий и их участие в хемотаксисе
Добавил пользователь Евгений Кузнецов Обновлено: 22.01.2026
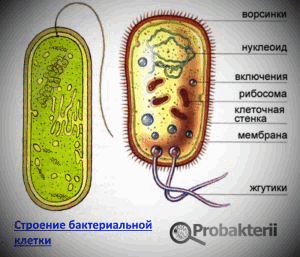
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
Обозначения:
Клеточная стенка
В клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка).
В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид (ЛПС). Липополисахарид наружной мембраны состоит из трех фрагментов: липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий; ядра, или стержневой, коровой части (лат. core — ядро), относительно консервативной олигосахаридной структуры (наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота); высоковариабельнои О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями (О-антиген). Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима,
пенициллина, защитных факторов организма образуются клетки с измененной (часто шаровидной) формой: протопласты — бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. Бактерии сферо- или протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами.
Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, «возвращаясь» в исходную бактериальную клетку.
Между наружной и цитоплазматической мембранами находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактомазы) и компоненты транспортных систем.
Цитоплазматическая мембрана
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым - промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами.
Цитоплазма
Цитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) - консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они являются запасными веществами для питания и энергетических потребностей бактерий. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид
Нуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК.
Кроме нуклеоида, представленного одной хромосомой, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды, представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь
Капсула - слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слиэь - мукоидные экзополисахариды, не имеющие четких границ. Слизь растворима в воде.
Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам), их еще называют гликокаликсом. Кроме синтеза
экзополисахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны.
Жгутики
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий
с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
Споры, капсулы, ворсинки, жгутики. Их строение, химический состав, функции, методы выявления.
Капсула или слизистый слой окружает оболочку ряда бактерий. Выделяют микрокапсулу, выявляемую при электронной микроскопии в виде слоя микрофибрилл, и макрокапсулу, обнаруживаемую при световой микроскопии. Капсула является защитной структурой (прежде всего от высыхания), у ряда микробов- фактором патогенности, препятствует фагоцитозу, ингибирует первые этапы защитных реакций- распознавание и поглощение. У сапрофитов капсулы образуются во внешней среде, у патогенов- чаще в организме хозяина. Существут ряд методов окраски капсул в зависимости от их химического состава. Капсула чаще состоит из полисахаридов (наиболее распространенная окраска- по Гинсу), реже- из полипептидов.
Жгутики. Подвижные бактерии могут быть скользящие (передвигаются по твердой поверхности в результате волнообразных сокращений) или плавающие, передвигающиеся за счет нитевидных спирально изогнутых белковых (флагеллиновых по химическому составу) образований- жгутиков.
По расположению и количеству жгутиков выделяют ряд форм бактерий.
1.Монотрихи- имеют один полярный жгутик.
2.Лофотрихи- имеют полярно расположенный пучок жгутиков.
3.Амфитрихи- имеют жгутики по диаметрально противоположным полюсам.
4.Перитрихи- имеют жгутики по всему периметру бактериальной клетки.
Способность к целенаправленному движению (хемотаксис, аэротаксис, фототаксис) у бактерий генетически детерминирована.
Фимбрии или реснички - короткие нити, в большом количестве окружающую бактериальную клетку, с помощью которых бактерии прокрепляются к субстратам (например, к поверхности слизистых оболочек). Таким образом, фимбрии являются факторами адгезии и колонизации.
F- пили (фактор фертильности) - аппарат конъюгации бактерий, встречаются в небольшом количестве в виде тонких белковых ворсинок.
Эндоспоры и спорообразование.
Спорообразование- способ сохранения определенных видов бактерий в неблагоприятных условиях среды. Эндоспоры образуются в цитоплазме, представляют собой клетки с низкой метаболической активностью и высокой устойчивостью (резистентностью) к высушиванию, действию химических факторов, высокой температуры и других неблагоплиятных факторов окружающей среды. При световой микроскопии часто используют метод выявления спор по Ожешко. Высокая резистентность связана с большим содержанием кальциевой соли дипиколиновой кислоты в оболочке спор. Расположение и размеры спор у различных микроорганизмов отличается, что имеет дифференциально- диагностическое (таксономическое) значение. Основные фазы “жизненного цикла” спор- споруляция (включает подготовительную стадию, стадию предспоры, образования оболочки, созревания и покоя) и прорастание, заканчивающееся образованием вегетативной формы. Процесс спорообразования генетически обусловлен.
Некультивируемые формы бактерий.
У многих видов грамотрицательных бактерий, не образующих спор, существует особое приспособительное состояние- некультивируемые формы. Они обладают низкой метаболической активностью и активно не размножаются, т.е. не образуют колоний на плотных питательных средах, при посевах не выявляются. Обладают высокой устойчивостью и могут сохранять жизнеспособность в течение нескольких лет. Не выявляются классическими бактериологическими методами, обнаруживаются только при помощи генетических методов ( полимеразной цепной реакции- ПЦР).
Жгутики и движение бактерий
Критерий, используемый для оценки отчета об исследовании; конкретно в отношении того, логична и информационно точна ли аргументация отчета.
(отчет может быть неточным: малоответственная обработка данных, нелогичность аргументации, неумелое построение фраз).
Ясность
Критерий, используемый для оценки отчета; конкретно в отношении того, точна ли фразеология отчета.
Применять простое правило:
Отчет должен быть строго организован:
- определить цель отчета;
- общий набросок главных акцентов;
- расставить акценты в логическом порядке;
- определить предмет исследования.
(убрать жаргон, использовать короткие слова, писать просто и естественно, избегать нечетких определений)
Критерий, используемый для оценки того, насколько живо и непосредственно написан отчет
(прочитать отчет вслух)
- титульный лист
- содержание
- краткий обзор
- введение
- основная часть
- заключительные рекомендации
- приложение
Большинство бактерий передвигаются при помощи жгутиков. Рассмотреть жгутики можно только в электронном микроскопе. В световом микроскопе без специальных методов обработки отдельные жгутики не видны.
По расположению и числу жгутиков на поверхности клетки бактерии подразделяются:
• на монотрихи - имеют один жгутик (например, бактерии родов Caulobacter и Vibrio);
• лофотрихи - имеют на одном или на обоих полюсах клетки пучок жгутиков (например, бактерии родов Pseudomonas, Chromatium);
• амфитрихи - имеют по жгутику на обоих полюсах клетки (например, бактерии рода Spirillum);
• перитрихи - большое количество жгутиков, располагающихся по всей поверхности клетки (например, бактерии вида E.coli и рода Erwinia) (рис. 1).
Жгутики представляют собой спирально закрученные нити, состоящие из специфического белка флагеллина. Флагеллин построен из субъединиц с относительно малой молекулярной массой. Субъединицы располагаются по спирали вокруг внутреннего свободного пространства. Аминокислотный состав флагеллина у разных видов бактерий может варьировать.
Рисунок 1 - Типы жгутикования у бактерий
Жгутик состоит из трех частей: нити, крюка и базального тельца (рис. 2). С помощью базального тельца, в которое входит центральный стержень и кольца, жгутик закреплен в цитоплазматической мембране и клеточной стенке. Количество колец у грамотрицательных и грамположительных бактерий различно. У грамотрицательных бактерий имеются четыре кольца: L, P, S, M. Из них L и P - наружная пара колец; S и M внутренняя пара колец. L-кольцо закреплено в наружной мембране, P - в пептидогликановом слое клеточной стенки, S - в периплазматическом пространстве, а M - в цитоплазматической мембране. У грамположительных бактерий базальное тельце устроено проще. Оно состоит только из двух колец: S и M, т. е. только из внутренней пары колец, которые размещаются в цитоплазматической мембране и клеточной стенке.
Жгутики бактерий по характеру работы подобны корабельному винту. Если клетка имеет много жгутиков, они при движении собираются в пучок, который образует своеобразный пропеллер. Пучок жгутиков, быстро вращаясь против часовой стрелки, создает силу, заставляющую бактерию двигаться почти по прямой линии. После того как направление вращения жгутиков изменяется, пучок расплетается и клетка останавливается, вместо поступательного движения она начинает хаотически вращаться, ее ориентация изменяется. В тот момент, когда все жгутики бактерии снова начнут синхронно вращаться против часовой стрелки, образовав пропеллер, толкающий бактерию, направление ее поступательного движения будет отличаться от первоначального. Таким способом бактерия может изменять направление своего движения.
Рисунок 2 - Структура жгутика
Так как у грамположительных бактерий наружная пара колец отсутствует, то считают, что для вращения жгутиков необходимо наличие только внутренней пары (кольца S и M). Эти кольца, соединенные с вращающимся стержнем, выступающим наружу, и образуют так называемый электромотор, обеспечивающий движение жгутика (рис. 2). На периферии кольца М находятся белки MotB. Белки MotА встроены в цитоплазматическую мембрану и примыкают к краям колец M и S. Вращающий момент возникает за счет взаимодействия субъединиц белка MotВ с белковыми субъединицами MotА. В белковых субъединицах MotА имеется по два протонных полуканала. Через эти протонные полуканалы переносятся протоны из периплазматического пространства в цитоплазму бактерий (подобно протонному каналу АТФ-синтазы). В результате переноса протонов через белки MotА и MotВ происходит вращение кольца М. Установлено, что один полный оборот кольца М связан с переносом через мембрану около 1000 протонов. Таким образом, в качестве источника энергии для вращения жгутиков используется протондвижущая сила, возникающая в цитоплазматической мембране.
Для подвижных бактерий характерны таксисы, т. е. направленная двигательная реакция в ответ на определенный фактор. В зависимости от природы различают хемотаксис, фототаксис, магнитотаксис и вискозитаксис.
Хемотаксис - движение бактерий относительно источника химического вещества. Для каждого микроорганизма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы, или эффекторы. Среди эффекторов выделяют: аттрактанты - вещества, которые притягивают бактерии; репелленты - вещества, которые отпугивают бактерии.
Фототаксис - движение к источнику света или от него, свойственное фототрофным бактериям.
Магнитотаксис - способность бактерий передвигаться по силовым линиям магнитного поля Земли или магнита. Выявлен в клетках бактерий, содержащих магнитосомы и распространенных в водных экосистемах разного типа.
У ряда бактерий выявлен вискозитаксис - способность реагировать на изменение вязкости раствора и передвигаться в направлении ее увеличения или уменьшения. За чувствительность бактерий к градиенту концентраций определенных факторов ответственны специфические рецепторы. Рецептор реагирует на эффектор и передает сигнал определенного типа на базальное тельце жгутика.
Ворсинки (или фимбрии)
Ворсинки, или фимбрии, - поверхностные структуры, которые состоят из белка пилина и не выполняют функцию движения. По размерам они короче и тоньше жгутиков. Число фимбрий на поверхности клетки колеблется от 1-2 до нескольких тысяч, их имеют как кокковидные, так и палочковидные бактерии. Различают два типа фимбрий: общие и специфические.
Фимбрии общего типа выполняют функцию прикрепления клетки к поверхности субстрата. Не исключается возможность их участия в поступлении крупномолекулярных соединений в цитоплазму клетки.
Специфические ворсинки - половые пили, обнаруженные у клеток так называемых доноров, т. е. у клеток, содержащих половой фактор (F-плазмиду) или другие донорспецифические плазмиды. Если в клетке бактерий находится половой фактор, то на их поверхности синтезируются одна-две половые F-пили на клетку. Они имеют вид полых белковых трубочек длиной от 0,5 до 10 мкм. F-пили играют определяющую роль в образовании конъюгационных пар при переносе генетического материала от клетки донора в клетку реципиента.
Это важно знать:
Этапы жизненного цикла организации Жизненный цикл организации - ее предсказуемые изменения с определенной последовательностью состояний в течение времени.
Примеры решения задач. Определите рентабельность продукции по следующим данным: количество выпущенных изделий за квартал - 1 500 штук Задача 1 Определите рентабельность продукции по следующим данным: количество выпущенных изделий за квартал - 1 500 штук.
Общее заключение по уроку, выводы и предложения 1. Общая оценка решения поставленных задач: в процессе урока решались образовательные.
Модели и стратегии поведения личности в конфликте Анализ конфликта и выбор адекватных решений по управлению им обуславливает необходимость учета типичные моделей поведения.
Виды технического обслуживания и ремонта вагонов На железных дорогах действует планово-предупредительная система ремонта вагонов.
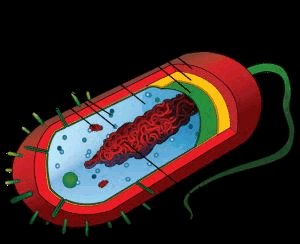
Строение бактериальной клетки.
Обязательными органоидами являются: ядерный аппарат, цитоплазма, цитоплазматическая мембрана.
Необязательными (второстепенными) структурными элементами являются: клеточная стенка, капсула, споры, пили, жгутики.
1.В центре бактериальной клетки находится нуклеоид- ядерное образование, представленное чаще всего одной хромосомой кольцевидной формы. Состоит из двухцепочечной нити ДНК. Нуклеоид не отделен от цитоплазмы ядерной мембраной.
2.Цитоплазма- сложная коллоидная система, содержащая различные включения метаболического происхождения (зерна волютина, гликогена, гранулезы и др.), рибосомы и другие элементы белоксинтезирующей системы, плазмиды (вненуклеоидное ДНК), мезосомы (образуются в результате инвагинации цитоплазматической мембраны в цитоплазму, участвуют в энергетическом обмене, спорообразовании, формировании межклеточной перегородки при делении).
3.Цитоплазматическая мембрана ограничивает с наружной стороны цитоплазму, имеет трехслойное строение и выполняет ряд важнейших функций- барьерную (создает и поддерживает осмотическое давление), энергетическую (содержит многие ферментные системы- дыхательные, окислительно- восстановительные, осуществляет перенос электронов), транспортную (перенос различных веществ в клетку и из клетки).
4.Клеточная стенка- присуща большинству бактерий (кроме микоплазм, ахолеплазм и некоторых других не имеющих истинной клеточной стенки микроорганизмов). Она обладает рядом функций, прежде всего обеспечивает механическую защиту и постоянную форму клеток, с ее наличием в значительной степени связаны антигенные свойства бактерий. В составе - два основных слоя, из которых наружный- более пластичный, внутренний- ригидный.
Основное химическое соединение клеточной стенки, которое специфично только для бактерий- пептидогликан (муреиновые кислоты). От структуры и химического состава клеточной стенки бактерий зависит важный для систематики признак бактерий- отношение к окраске по Граму. В соответствии с ним выделяют две большие группы- грамположительные (“грам+”) и грамотрицательные (“грам - “) бактерии. Стенка грамположительных бактерий после окраски по Граму сохраняет комплекс йода с генциановым фиолетовым (окрашены в сине- фиолетовый цвет), грамотрицательные бактерии теряют этот комплекс и соответствующий цвет после обработки и окрашены в розовый цвет за счет докрашивания фуксином.
Особенности клеточной стенки грамположительных бактерий.
Мощная, толстая, несложно организованная клеточная стенка, в составе которой преобладают пептидогликан и тейхоевые кислоты, нет липополисахаридов (ЛПС), часто нет диаминопимелиновой кислоты.
Особенности клеточной стенки грамотрицательных бактерий.
Клеточная стенка значительно тоньше, чем у грамположительных бактерий, содержит ЛПС, липопротеины, фосфолипиды, диаминопимелиновую кислоту. Устроена более сложно- имеется внешняя мембрана, поэтому клеточная стенка трехслойная.
При обработке грамположительных бактерий ферментами, разрушающими пептидогликан, возникают полностью лишенные клеточной стенки структуры- протопласты. Обработка грамотрицательных бактерий лизоцимом разрушает только слой пептидогликана, не разрушая полностью внешней мембраны; такие структуры называют сферопластами. Протопласты и сферопласты имеют сферическую форму (это свойство связано с осмотическим давлением и характерно для всех безклеточных форм бактерий).
L- формы бактерий.
Под действием ряда факторов, неблагоприятно действующих на бактериальную клетку (антибиотики, ферменты, антитела и др.), происходит L- трансформация бактерий, приводящая к постоянной или временной утрате клеточной стенки. L- трансформация является не только формой изменчивости, но и приспособления бактерий к неблагоприятным условиям существования. В результате изменения антигенных свойств (утрата О- и К- антигенов), снижения вирулентности и других факторов L- формы приобретают способность длительно находиться (персистировать) в организме хозяина, поддерживая вяло текущий инфекционный процесс. Утрата клеточной стенки делает L- формы нечувствительными к антибиотикам, антителам и различным химиопрепаратам, точкой приложения которых является бактериальная клеточная стенка. Нестабильные L- формы способны реверсировать в классические (исходные) формы бактерий, имеющие клеточную стенку. Имеются также стабильные L- формы бактерий, отсутствие клеточной стенки и неспособность реверстровать которых в классические формы бактерий закреплены генетически. Они по ряду признаков очень напоминают микоплазмы и другие молликуты- бактерии, у которых клеточная стенка отсутствует как таксономический признак. Микроорганизмы, относящиеся к микоплазмам- самые мелкие прокариоты, не имеют клеточной стенки и как все бактериальные бесстеночные структуры имеют сферическую форму.
К поверхностным структурам бактерий (необязательным, как и клеточная стенка), относятся капсула, жгутики, микроворсинки.
Значение пилей в жизни бактерий
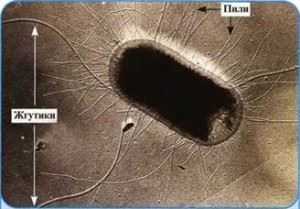
Наряду со жгутиками прокариоты могут обладать и другими внеклеточными образованиями. В середине ХХ века было установлено, что бактерии способны формировать специфическую группу поверхностных образований. Их называли ворсинками, ресничками, фимбриями. Сегодня их называют пилями бактерий.
Внешний вид фимбрий
Внешне пили, или фимбрии, как их называли до 1956 г., выглядят как микроскопические волоски, покрывающие клетку бактерии. На 1 клетку прокариота может приходиться от нескольких единиц до тысяч ворсинок.
Хотя они, как и жгутики, являются поверхностными образованиями, однако между собой имеют больше различий, чем сходств.
По размеру пили намного меньше жгутиков, в среднем в 3 раза тоньше (не более 10 нм), и длиной не превышают 1,5 мкм.
По строению, несмотря на то, что как пили, так и жгутики состоят из белковых клеток, они также различаются:
- пили, или фимбрии, представляют собой легкую цепочку проводящих белков цилиндрической формы, отходящую от поверхностного слоя клетки;
- жгутики являются более громоздкими по строению, с наличием сложных структур (стержень, базальное тело, кольца и другое).
Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Для чего прокариотам пили
К примеру, если жгутики бактерий обеспечивают возможность передвигаться, то фимбрии не имеют никакого отношения к перемещению в пространстве и присутствуют как у движущихся, так и у неподвижных бактерий.
Неподтвержденным исключением можно считать участие пилей 4 типа в скользящем движении прокариотов всей колонией, характерном для многих протеобактерий (и миксобактерий), скользящих бактерий, циано-, несерных бактерий и других.
В отличие от жгутиков функции пилей бактерий изучены достаточно слабо, но совершенно очевидно, что одной из них является способность обеспечить прикрепление клетки бактерии к питательному субстрату.
Разные типы ворсинок
Пили не являются однородными образованиями, их различают как минимум 4 типа, каждый из которых выполняет свои функции, причем одна клетка может являться носителем несколько разных типов фимбрий.
Пили 1 типа
Фимбрии бактерий 1 типа образуются из пилина (белок) и отличаются крайне прочной связью с прокариотом. Чтобы отделить такую фимбрию от бактериальной клетки, требуются усилия, многократно превышающие необходимое воздействие для отделения половых пилей или жгутиков.
Для пилей 1 типа характерно расположение перитрихиально - по всей поверхности бактерии.
Исследования методами выявления свойств показали, что пили 1 типа являются химически устойчивыми образованиями - они инертны к растворам щелочей, мочевине и трипсину (фермент, расщепляющий белки).
Разрушаются пили 1 типа при кипячении в растворах с высокой кислотностью, при этом методе воздействия происходит необратимое разрушение (денатурация) белка, образующего фимбрию.
Характерной особенностью пилей 1 типа является:
- способность образовывать пленки и придавать бактериям гидрофобные свойства;
- способность вызывать агглютинацию эритроцитов (выпадение в осадок в результате склеивания) под действием агглютининов.
Основными функциями являются:
- адгезивная - прикрепление бактерий к субстратам;
- защитная - объединение клеток прокариотов, получивших гидрофобные свойства, в группы;
- участие в процессах метаболизма клетки - увеличение всасывающей поверхности.
Ворсинки 2 типа
Эта группа имеет очень много общего с предыдущей, однако не обладает характерными особенностями 1 типа - пили не участвуют в формировании пленок и не приклеиваются к эритроцитам (агглютинация), провоцируя выпадение их в осадок.
Столь близкое сходство позволяет предположить, что пили 2 типа являются мутантной формой 1 типа.
Половые фимбрии (3 тип)
Современные методы выявления позволили определить, что при горизонтальном переносе генетического материала (конъюгации) ключевую роль играют половые пили.
Возможность непосредственного контакта двух бактериальных клеток с последующей конъюгацией была выявлена в 50-х годах прошлого века в результате исследований двух американских биохимиков - Д.Ледербейга и Э.Тейтема. Данный процесс имеет большое практическое значение, так как позволяет производить обмен наследственными признаками организмам, размножающимся только прямым делением.
Половые фимбрии, их называют F-пили, присутствуют только у бактериальных штаммов, обладающих фактором трансмиссивности - это может быть автономный репликон или его часть.
F-пили представляют собой цилиндрические белковые образования большего диаметра, чем пили 1 или 2 типа, расположенные перпендикулярно к поверхности.
В случае потери F-пили восстанавливаются - в течение 30 секунд пиль достигает половины своей величины. Для формирования полноценной трубочки необходимо от 4 до 5 минут.
Современные методы выявления позволили определить, что на поверхности бактерии F-пили сохраняются в течение 5 минут, после чего сбрасываются, и процесс повторяется.
F-пили значительно отличаются от ворсинок 1 и 2 типа как по строению, так и по свойствам.
В отличие от последних F-пили легко отделяются от бактериальной клетки даже при незначительном встряхивании.
Методами физико-химического анализа было определено, что в составе F-пили отсутствует целый ряд α-аминокислот, характерных для белка пилей 1 типа, но через ковалентную связь присоединены остаток D-глюкозы и две фосфатные группы.
В связи с иной химической структурой на F-пили не адсорбируются обычные фаги, а только специфичные для них, называемые мужскими фагами.
Участие F-пили в процессе передачи информации
Процесс передачи части генетической информации подразумевает наличие пары клетка-донор и клетка-реципиент.
- Первоначально клетка-донор формирует F-пиль.
- F-пиль донора фиксируется на клетке-реципиенте.
- В F-плазмиде донорной клетки осуществляется разрыв одной нити ДНК, которая передается реципиенту.
- Обе бактерии достраивают вторую цепочку ДНК и восстанавливают F- плазмиду. Клетка-реципиент превращается в донора.
Микроскопические методы исследований позволили определить, что образование F-пилей характерно только для растущих и активных клеток, при переходе в стационарную фазу роста бактерии теряют свою способность образовывать половые пили и становятся плохими донорами.
Специфическая направленность фимбрий 4 типа
Пили 4 типа принимают участие в обеспечении разновидности скользящего движения бактерий всей колонией.
Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
- А-система - секретирует слизь на полюсе направления движения микроорганизма.
- S-система - роение; обеспечивается последовательным сокращением и удлинением пилей 4 типа, подобное подтягиванию.
Механизм данного вида бактериального движения на сегодняшний день находится в процессе изучения, и большинство выводов носят предположительный характер.
Образование высшее филологическое. В копирайтинге с 2012 г., также занимаюсь редактированием/размещением статей. Увлечения — психология и кулинария.
Читайте также: